Abstract
The pathogenesis and pathophysiology of Acanthamoeba infections remain incompletely understood. Phos-pholipases are known to cleave phospholipids, suggesting their possible involvement in the host cell plasma membrane disruption leading to host cell penetration and lysis. The aims of the present study were to determine phospholipase activities in Acanthamoeba and to determine their roles in the pathogenesis of Acanthamoeba. Using an encephalitis isolate (T1 genotype), a keratitis isolate (T4 genotype), and an environmental isolate (T7 genotype), we demonstrated that Acanthamoeba exhibited phospholipase A2 (PLA2) and phospholipase D (PLD) activities in a spectrophotometry-based assay. Interestingly, the encephalitis isolates of Acanthamoeba exhibited higher phospholipase activities as compared with the keratitis isolates, but the environmental isolates exhibited the highest phospholipase activities. Moreover, Acanthamoeba isolates exhibited higher PLD activities compared with the PLA2. Acanthamoeba exhibited optimal phospholipase activities at 37℃ and at neutral pH indicating their physiological relevance. The functional role of phospholipases was determined by in vitro assays using human brain microvascular endothelial cells (HBMEC), which constitute the blood-brain barrier. We observed that a PLD-specific inhibitor, i.e., compound 48/80, partially inhibited Acanthamoeba encephalitis isolate cytotoxicity of the host cells, while PLA2-specific inhibitor, i.e., cytidine 5'-diphosphocholine, had no effect on parasite-mediated HBMEC cytotoxicity. Overall, the T7 exhibited higher phospholipase activities as compared to the T4. In contract, the T7 exhibited minimal binding to, or cytotoxicity of, HBMEC.
-
Key words: Acanthamoeba, human brain microvascular endothelial cell, phospholipase A2, phospholipase D, adhesion, cytotoxicity
INTRODUCTION
Acanthamoeba is a free-living protozoan and has been isolated from diverse environments, including air, soil, tap water, or swimming pools [
1]. It is recognized as one of the most ubiquitous protozoans [
1].
Acanthamoeba is an opportunistic human pathogen and can cause a rare form of granulomatous encephalitis which almost always results in death or a common keratitis with consequences of blindness [
2-
4]. However, the pathogenesis and pathophysiology associated with
Acanthamoeba infections remain incompletely understood.
Acanthamoeba has been shown to demonstrate several extracellular toxins, particularly extracellular proteases which might contribute towards its pathogenicity. Previous studies have shown the presence of phospholipases in
Acanthamoeba; however, our knowledge of the role of phospholipases in the virulence of
Acanthamoeba is unclear [
5-
7]. Besides their role in remodelling of the plasma membrane of the parasite itself, based on the fact that phospholipases are known to cleave phospholipids, they may involve in the host cell plasma membrane disruptions leading to penetration of the host cells, and cell lysis [
5-
7]. This suggests that an understanding of the phospholipases in the biology and pathogenesis of
Acanthamoeba may identify targets for therapeutic interventions.
Phospholipases are a diverse group of enzymes that hydrolyze ester linkage in glycerophospholipids and are involved in the biosynthesis and degradation of membrane lipids. Thus, the activities of phospholipases can result in membrane dysfunction. The 5 major known phospholipases are A
1, A
2, B, C, and D, each having the ability to cleave a specific ester bond in the phospholipid substrate of the target membrane [
8]. For example, PLA
1 hydrolyses the fatty acyl ester bond at the
sn-1 position of the glycerol moiety, while PLA
2 removes the fatty acid at the
sn-2 position of glycerol [
8]. The results of both the PLA
1 and PLA
2 action are free fatty acids or acyl lysophospholipid. The latter are further cleaved by the action of lysophospholi pases. Phospholipase B has both hydrolase activities, such as cleaving fatty acids from phospholipids and lysophospholipids as well as acyltransferase activity, for instance, the production of phospholipids by transferring a free fatty acid to a lysophospholipid [
8]. Phospholipase C hydrolyses phosphodiester bond to yield 1,2-diacylglycerol depending on the phospholipid class involved, while PLD hydrolyses the phosphodiester bond between the phosphate and the polar head group, producing phosphatidic acid and choline [
8-
10]. All phospholipases are present in multiple forms. For example, approximately 20 different PLA
2 enzymes have been identified in mammalian tissues and have been broadly classified into 3 families based on their calcium requirement for catalytic activity [
8,
11,
12]. Different parts of the breakages by PLA
2 and PLD are focused on this study. Victoria and Korn [
6] observed that PLA of
Acanthamoeba castellanii was located in the plasma membrane [
6]. It may be affected on binding of
Acanthamoeba to target cells. The aims of the present study were to determine PLA
2 and PLD activities in
Acanthamoeba and to determine their role in
Acanthamoeba binding to, and cytotoxicity of, human brain microvascular endothelial cells (HBMEC).
MATERIALS AND METHODS
Culture of Acanthamoeba
All chemicals were purchased from Sigma Laboratories (Poole, Dorset, UK), unless otherwise stated. The following
Acanthamoeba isolates of different pathogenicity were used; (1) a clinical isolate of
A. castellanii belonging to T4 genotype, isolated from a keratitis patient (ATCC 50492) (American Type Culture Collection, Manassas, USA), (2) a clinical isolate of
A. castellanii belonging to T1 genotype, isolated from an encephalitis patient (ATCC 50494), and (3) an environmental isolate of
Aanthamoeba astronyxis belonging to T7 genotype, isolated from soil (ATCC 30137). All amoebae were grown in 15 ml of PYG medium (proteose peptone 0.75% w/v, yeast extract 0.75% w/v, and glucose 1.5% w/v) at 30℃ and the media was refreshed 17-20 hr prior to experiments as previously described [
13]. This resulted in more than 99% amoebae in trophozoite forms, which were used for subsequent experiments.
The primary HBMEC were isolated from the human brain tissue and cultured as previously described [
14,
15]. In brief, HBMEC were purified by fluorescent activated cell sorting (FACS) and tested for endothelial characteristics, such as the expression of endothelial markers, F-VIII, carbonic anhydrase IV, and the uptake of acetylated low density lipoprotein (AcLDL), resulting in more than 99% pure cultures. HBMEC were routinely grown on rat tail collagen-coated dishes in RPMI-1640 containing 10% heat inactivated fetal bovine serum, 10% Nu serum, 2 mM glutamine, 1 mM sodium pyruvate, penicillin (100 U/ml), streptomycin (100 U/ml), non-essential amino acids, and vitamins [
14,
15].
Adhesion assays were performed as previously described [
4]. Briefly, HBMEC were grown to confluency in 24-well plates.
Acanthamoeba isolates (2×10
5 amoebae/0.5 ml/well) were in-cubated with cell monolayers in RPMI-1640 containing 2 mM glutamine, 1 mM sodium pyruvate, and non-essential amino acids. The plates were incubated at 37℃ in a 5% CO
2 incubator. After 60 min incubations, the numbers of unbound amoebae in the supernatants were determined by hemocytometer counting. The percentage of bound amoebae was calculated as follows: Number of unbound amoebae/Total number of amoebae×100=% unbound amoebae. The numbers of bound amoebae were deduced as follows: 100-% unbound amoebae=% bound amoebae.
To determine the role of PLA2 and PLD in Acanthamoeba adhesion to the HBMEC, assays were performed in the presence of phospholipase inhibitors. Briefly, amoebae were pre-incu-bated with the phospholipase inhibitors for 45 min at room temperature. Following this incubation, amoebae plus inhibitors were transferred to HBMEC monolayers for adhesion assays as described above. The following phospholipase inhibitors were used: cytidine 5'-diphosphocholine, i.e., PLA2 inhibitor; and compound 48/80, i.e., PLC/PLD inhibitor.
Cytotoxicity assay
To determine the role of phospholipases in
Acanthamoeba-mediated HBMEC death, cytotoxicity assays were performed as previously described [
13]. In brief, amoebae in the presence or absence of phospholipase inhibitors were added to HBMEC monolayers as described for adhesion assays. The plates were observed periodically for monolayer disruptions under a phase-contrast microscope for up to 24 hr. Following this incubation, the supernatants were collected and examined for host cell cytotoxicity by measuring lactate dehydrogenase (LDH) release (cytotoxicity detection kit; Roche Applied Science, Lewes, East Sussex, UK). In brief, the supernatants of co-cultures of
Acanthamoeba and the host cells were assessed for the presence of LDH, the release of which is considered as an estimate of cell death. The percentage LDH release was calculated as follows: (LDH activity in experimental sample as measured by optical density at 492 nm-LDH activity in control samples]/(total LDH activity release-LDH activity in control samples)×100=% cy-totoxicity. Control samples were obtained from host cells or
Acanthamoeba incubated alone. Total LDH activity release was determined by total host cell lysis with 1% Triton X-100 for 30 min at 37℃.
To determine the activity of phospholipases, the characteristics of L-α-glycerophosphate oxidase for PLA
2 and choline oxidase for PLD were used.
Acanthamoeba PLA
2 activities were determined using parasite lysates and their conditioned medium in a spectrophotometric-based assay. To prepare lysates, amoebae were lysed using freeze-thaw method as previously described [
16]. In brief, various numbers of amoebae were resuspended in 100 µl of PBS, followed by freeze-thaw for 3 times. To prepare the conditioned medium,
Acanthamoeba were incubated in RPMI-1640 for 24 hr. Next day, the cell-free conditioned medium was collected by centrifugation and used for PLA
2 assays. A 100 µl of amoebae lysate or conditioned medium was incubated with 900 µl of PLA
2 substrate solution (0.2 M D, L-α-glycerophosphate, 0.1% 4-aminoantipyrine, 0.1% phenol, and 0.025% peroxidase dissolved in 0.125 M Tris-HCl buffer, pH 8.0). The reaction mixtures were incubated for 2 hr. After this incubation, the mixtures were centrifuged at 900
g for 5 min and supernatants were collected and their optical density determined at 500 nm. The PLA
2 activity was calculated as follows: (OD test-OD blank)×Vt×df/13.3×0.5×t×1.0×Vs=enzymatic activity (units per ml). Here, Vt is total volume; Vs is sample volume; 13.3 is the millimolar extinction coefficient of quinoneimine dye under the assay condition; 0.5 is the factor based on the fact that 1 mole of H
2O
2 produces half a mole of quinoneimine dye; t is reaction time; 1.0 is light path (cm); and df is the dilution factor. To determine the optimal activity, PLA
2 assays were performed using various numbers of amoebae at different pH and temperatures.
Acanthamoeba PLD activities were determined using parasite lysates and their conditioned medium in a spectrophotometric-based assay. The lysates and conditioned medium were prepared as described above. A 100 µl of amoebae lysate or conditioned medium was incubated with 900 µl of PLD substrate solution (2.1% choline chloride, 1% 4-aminoantipyrine, 1% phenol, and 5 mg peroxidase in 0.1 M Tris-HCl buffer, pH 8.0). The reaction mixtures were incubated for 2 hr. After this incubation, the mixtures were centrifuged at 900 g for 5 min, and supernatants were collected and their optical density de-termined at 500 nm. The PLD activity was calculated as follows: (OD test-OD blank)/min×Vt×df/12.0×0.5×1.0×Vs=enzymatic activity (units per ml). Here, Vt is total volume; Vs is sample volume; 12.0 is the millimolar extinction coefficient of quinoneimine dye under the assay condition; 0.5 is the factor based on the fact that 1 mole of H2O2 produces half a mole of quinoneimine dye; t is reaction time; 1.0 is light path (cm); and df is the dilution factor.
RESULTS
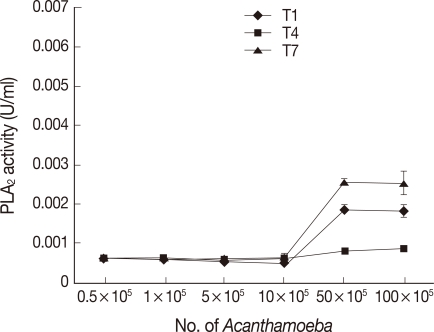
Acanthamoeba exhibit PLA2 activities
Both clinical and environmental isolates of
Acanthamoeba exhibited PLA
2 activities (
Fig. 1). As expected, optimal PLA
2 activities were observed with increasing numbers of
Acanthamoeba. Interestingly, the environmental isolate belonging to T7 genotype exhibited the maximum PLA
2 activities, followed by the encephalitis isolate belonging to the T1 genotype, while the keratitis isolate belonging to the T4 genotype exhibited the least PLA
2 activities (
Fig. 1). Of interest,
Acanthamoeba conditioned medium did not exhibit PLA
2 activities in the assay conditions tested in the present study, suggesting that PLA
2 are cell-associated and/or not released (data not shown).
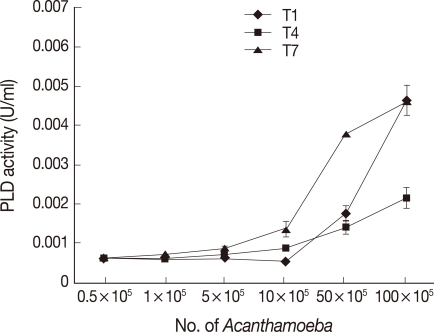
The results revealed that both clinical and environmental isolates of
Acanthamoeba exhibit PLD activities (
Fig. 2). Again, the encephalitis isolate (T1) and environmental isolate (T7) exhibited the highest levels of PLD activities compared with the keratitis isolate (T4) (
Fig. 2). It was interesting to note that PLD activities in
Acanthamoeba were significantly greater compared with
Acanthamoeba PLA
2 activities (
Figs. 1,
2). As for PLA
2, the
Acanthamoeba conditioned medium did not exhibit PLD activity in the assay conditions tested in the present study, indicating that PLD activities are cell-associated and/or not released (data not shown).
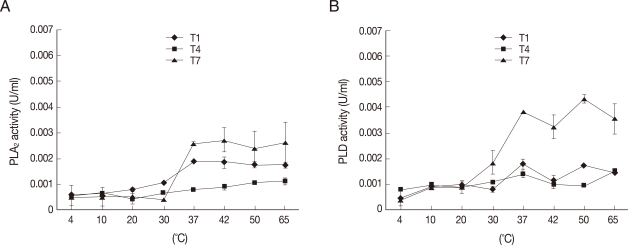
To determine the optimum phospholipase activities, assays were performed at various temperatures from 4℃ to 65℃. The optimal phospholipase activities were observed at 37℃ suggesting their physiological relevance (
Fig. 3). Similar findings were observed both with PLA
2 and PLD activities. Activities were also observed at 65℃, suggesting that
Acanthamoeba PLA
2 and PLD activities are stable at higher temperatures (
Fig. 3).
Next to determine the pH for optimal phospholipase activities, assays were performed at various pH. Our findings revealed that
Acanthamoeba exhibit optimal PLA
2 and PLD activities at neutral pH indicating their physiological relevance (
Fig. 4). Both clinical and environmental isolates exhibited optimal phospholipase activities at pH 7.5.
To determine whether phospholipases play a role in
Acanhamoeba binding to the host cells, adhesion assays were performed using the PLA
2 inhibitor, i.e., cytidine 5'-diphosphocholine, and the PLC/PLD inhibitor, i.e., compound 48/80. Our findings revealed that neither phospholipase inhibitor had any significant effects on the adhesion of clinical isolates of
Acanthamoeba to HBMEC (
Fig. 5). In contrast, adhesion of the environmental isolate of
Acanthamoeba was completely abolished in the presence of phospholipase inhibitors (
Fig. 5).
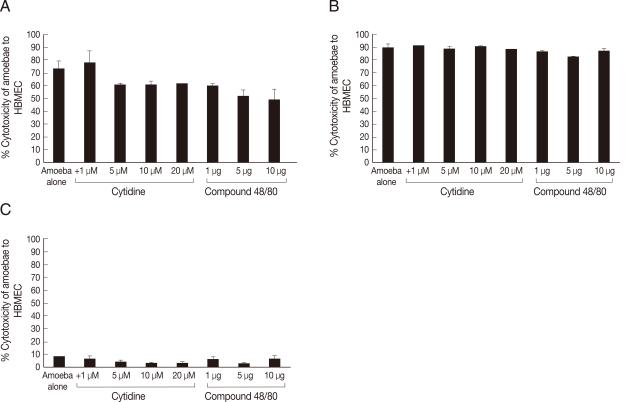
To determine the role of PLA
2 in
Acanthamoeba-mediated HBMEC death, cytotoxicity assays were performed in the presence of cytidine 5'-diphosphocholine, a PLA
2-specific inhibitor. The results revealed that cytidine 5'-diphosphocholine had no significant effect on parasite-mediated HBMEC cytotoxicity in all
Acanthamoeba isolates tested (
Fig. 6).
To determine whether PLC/PLD plays a role in
Acanthamoeba-mediated HBMEC death, cytotoxicity assays were performed in the presence of compound 48/80. Interestingly, compound 48/80 partially blocked the encephalitis isolate-mediated HBMEC cytotoxicity, i.e., 49% cell death in the presence of the inhibitor compared with 73% in the absence of the inhibitor (
Fig. 6). However, it had no effect on the keratitis isolate-mediated HBMEC cytotoxicity (
Fig. 6). In contrast, the effects of phospholipase inhibitors on the environmental isolate-mediated HBMEC cytotoxicity remained unclear. This was due to the fact that the environmental isolate exhibited minimal HBMEC cytotoxicity even in the absence of inhibitors.
DISCUSSION
Infections due to
Acanthamoeba have increased over the years. This is most likely due to the increasing numbers of contact lens wearers, increasing populations of immunocompromised patients, and global warming. This has led to a remarkable interest in the field of
Acanthamoeba pathogenesis and to design strategies to control and prevent
Acanthamoeba infections. In particular, over the last 2 decades, research in
Acanthamoeba virulence factors has gained significant attention to identify novel approaches for therapeutic interventions.
Acanthamoeba pathogenesis is attributed to their ability to bind to the host cells, induce host cell apoptosis, secrete proteases, exhibit ecto-ATPase activities, and switch phenotypes. The present study was undertaken to determine the roles of phospholipases (i.e., PLA
2 and PLD) in
Acanthamoeba and
Acanthamoeba-mediated binding to, and cytotoxicity of HBMEC in vitro. Our findings revealed that
Acanthamoeba show PLA
2 and PLD activities. This is consistent with previous studies that showed the presence of phospholipase activities in
Acanthamoeba [
5-
7].
It was interesting to note that the environmental isolate (belonging to the T7 genotype) exhibited higher phospholipase activities as compared to the keratitis isolate (belonging to the T4 genotype). In contrast, the environmental isolate exhibited minimal binding to or cytotoxicity of HBMEC. This is not a surprising finding. Being a free-living amoeba, the primary functions of phospholipases in
Acanthamoeba may be predominantly in the phospholipids turnover for membrane maintenance and remodelling. The fact that environmental isolate is significantly larger in size as compared to the keratitis isolate, thus larger plasma membranes, may explain its optimal PLA
2 and PLD activities, and thus rationalize these findings. In support, the size of keratitis isolate tested here is approximately 7 µm. The size of the encephalitis isolate is approximately 10 µm and the environmental isolate being the largest, approximately 17 µm. Another explanation may be that the environmental isolate exhibited distinct types or isoforms of phospholipases compared with the clinical isolates. For example, Sissons et al. [
17] have shown that both clinical and environmental isolates of
Acanthamoeba exhibit ecto-ATPase activities, however, clinical isolates exhibit distinct ecto-ATPases. This may be the case for phospholipases, which are normally required for routine cellular functions but pathogenic isolates may possess distinct phospholipases. The fact that compound 48/80 had no effect on keratitis isolate-mediated host cell death but partially inhibited encephalitis isolate-mediated HBMEC cytotoxicity further suggests possible differences in phospholipases in
Acanthamoeba belonging to different genotypes. Future studies are in progress to determine the molecular basis of these differences.
Of interest was the observation that compound 48/80, only partially inhibited encephalitis isolate-mediated HBMEC cytotoxicity. It is noteworthy that
Acanthamoeba pathogenesis is a multifactorial process involving adherence, phagocytosis, apoptosis, proteases, ecto-ATPases, and the inhibition of a single putative factor may not be sufficient to block their ability to produce host cell cytotoxicity. In support, previous studies have shown that inhibition of
Acanthamoeba binding to host cells is not adequate to block host cell cytotoxicity [
18]. At present, the inability of compound 48/80 to block keratitis isolate-mediated host cell death is unclear. To this end, the source of amoeba isolates, i.e., keratitis and encephalitis patients may be indicative of these differences. This possibly means that phospholipases from the encephalitis isolate may play a role in the pathogenesis of AGE. However, it is a subject which needs additional research and study.
The finding that phospholipase inhibitors did not clearly block
Acanthamoeba-mediated HBMEC cytotoxicity does not rule out their involvement in
Acanthamoeba pathogenesis. It is possible that cytotoxicity is a delayed event and that phospholipases are involved in the early events. Recent studies have shown that phospholipases may be involved in interference with the intracellular signalling pathways. For example, phospholipases generate lipids and lipid-derived products that act as mediators and second messengers, which may act as the modulators of signal transduction pathway [
19,
20]. Oishi et al. [
21] showed that lysophospholipids, by-end products of PLA
2 and PLB, induce activation of protein kinase C, which has diverse functions in host cell signalling pathways [
21]. Phospholipase C of
Clostridium perfringens induces expression of IL-8 synthesis in endothelial cells [
22,
23]. Phospholipase A facilitate host cell penetration of
Toxoplasma gondii [
24] and
Entamoeba histolytica [
25], while PLD has been shown to be an important virulence determinant of
Corynebacterium pseudotuberculosis in the persistence and spread of bacteria within the host [
26].
Overall, these studies suggest that
Acanthamoeba phospholipases may play a role in causing host cell damage or affect other cellular functions, such as inducing inflammatory responses and thus facilitating the virulence of
Acanthamoeba. The fact that phospholipases cleave phospholipids suggests their possible role in membrane disruptions and penetration into host cells. Future studies are needed to identify and characterize
Acanthamoeba phospholipases, which should help determine their potential role for therapeutic interventions and in differentiation of
Acanthamoeba isolates belonging to different genotypes. Some previous studies have shown that phospholipase C from
C. perfringens induced protection against
C. perfringens-mediated gas gangrene [
27]. In addition, targeting phospholipases using synthetic compounds are shown to have the potential to prevent
Candida infections [
28].
ACKNOWLEDGEMENTS
Funding for this paper was provided by Namseoul University.
References
- 1. Khan NA. Acanthamoeba: biology and increasing importance in human health. FEMS Microbiol Rev 2006;30:564-595.
- 2. Marciano-Cabral F, Cabral G. Acanthamoeba spp. as agents of disease in humans. Clin Microbiol Rev 2003;16:273-307.
- 3. Schuster FL, Visvesvara GS. Free-living amoebae as opportunistic and non-opportunistic pathogens of humans and animals. Int J Parasitol 2004;34:1001-1027.
- 4. Jung SY, Alsam S, Kim KS, Khan NA. Pathogen-pathogen interactions: a comparative study of Escherichia coli interactions with the clinical and environmental isolates of Acanthamoeba. World J Microbiol Biotechnol 2008;24:2339-2348.
- 5. Victoria EJ, Korn ED. Plasma membrane and soluble lysophospholipases of Acanthamoeba castellanii. Arch Biochem Biophys 1975;171:255-258.
- 6. Victoria EJ, Korn ED. Enzymes of phospholipid metabolism in the plasma membrane of Acanthamoeba castellanii. J Lipid Res 1975;16:54-60.
- 7. Cursons RTM, Brown TJ, Keys EA. Virulence of pathogenic free-living amoebae. J Parasitol 1978;64:744-745.
- 8. Ghannoum MA. Potential role of phospholipases in virulence and fungal pathogenesis. Clin Microbiol Rev 2000;13:122-143.
- 9. Exton JH. Phospholipase D-structure, regulation and function. Rev Physiol Biochem Pharmacol 2002;144:1-94.
- 10. McDermott M, Wakelam MJO, Morris AJ. Phospholipase D. Biochem Cell Biol 2004;82:225-253.
- 11. Akiba S, Sato T. Cellular function of calcium-independent phospholipase A2. Biol Pharm Bull 2004;27:1174-1178.
- 12. Kudo I. Diversity of phospholipase A2 enzymes. Foreword. Biol Pharm Bull 2004;27:1157.
- 13. Sissons J, Kim KS, Stins M, Jayasekera S, Alsam S, Khan NA. Acanthamoeba castellanii induces host cell death via a phosphatidylinositol 3-kinase-dependent mechanism. Infect Immun 2005;73:2704-2708.
- 14. Stins MF, Gilles F, Kim KS. Selective expression of adhesion molecules on human brain microvascular endothelial cells. J Neuroimmunol 1997;76:81-90.
- 15. Alsam S, Kim KS, Stins M, Rivas AO, Sissons J, Khan NA. Acanthamoeba interactions with human brain microvascular endothelial cells. Microb Pathog 2003;35:235-241.
- 16. Matin A, Stins M, Kim KS, Khan NA. Balamuthia mandrillaris exhibits metalloprotease activities. FEMS Immunol Med Microbiol 2006;47:83-91.
- 17. Sissons J, Alsam S, Jayasekera S, Khan NA. Ecto-ATPases of clinical and non-clinical isolates of Acanthamoeba. Microb Pathog 2004;37:231-239.
- 18. Leher H, Silvany R, Alizadeh H, Huang J, Niederkorn JY. Mannose induces the release of cytopathic factors from Acanthamoeba castellanii. Infect Immun 1998;66:5-10.
- 19. Dennis EA, Rhee SG, Billah MM, Hannun YA. Role of phospholipase in generating lipid second messengers in signal transduction. FASEB J 1991;5:2068-2077.
- 20. Serhan CN, Haeggström JZ, Leslie CC. Lipid mediator networks in cell signaling: update and impact of cytokines. FASEB J 1996;10:1147-1158.
- 21. Oishi K, Raynor RL, Charp PA, Kuo JF. Regulation of protein kinase C by lysophospholipids. Potential role in signal transduction. J Biol Chem 1988;263:6865-6871.
- 22. Bryant AE, Stevens DL. Phospholipase C and perfringolysin O from Clostridium perfringens upregulate endothelial cell-leukocyte adherence molecule 1 and intercellular leukocyte adherence molecule 1 expression and induce interleukin-8 synthesis in cultured human umbilical vein endothelial cells. Infect Immun 1996;64:358-362.
- 23. Bunting M, Lorant DE, Bryant AE, Zimmerman GA, McIntyre TM, Stevens DL, Prescott SM. Alpha toxin from Clostridium perfringens induces proinflammatory changes in endothelial cells. J Clin Invest 1997;100:565-574.
- 24. Saffer LD, Long Krug SA, Schwartzman JD. The role of phospholipase in host cell penetration by Toxoplasma gondii. Am J Trop Med Hyg 1989;40:145-149.
- 25. Long-Krug SA, Fischer KJ, Hysmith RM, Ravdin JI. Phospholipase A enzymes of Entamoeba histolytica: description and subcellular localization. J Infect Dis 1985;152:536-541.
- 26. McNamara PJ, Bradley GA, Songer JG. Targeted mutagenesis of the phospholipase D gene results in decreased virulence of Corynebacterium pseudotuberculosis. Mol Microbiol 1994;12:921-930.
- 27. Kameyama S, Sato H, Murata R. The role of alpha-toxin of Clostridium perfringens in experimental gas gangrene in guinea pigs. Jpn J Med Sci Biol 1972;25:200.
- 28. Hänel H, Kirsch R, Schmidts HL, Kottmann H. New systematically active antimycotics from the beta-blocker category. Mycoses 1995;38:251-264.
Fig. 1
Acanthamoeba exhibiting PLA2 activities. Both the encephalitis (A) and the environmental (C) isolates exhibited optimal, while the keratitis isolate (B) exhibited the least PLA2 activities. Results are representative of 3 independent experiments performed in triplicate. Bars represent standard errors.
Fig. 2Clinical and environmental Acanthamoeba isolates exhibiting PLD activities. As for PLA2, both the encephalitis (A) and the environmental (C) isolates exhibited optimal, while the keratitis isolate (B) exhibited the least PLD activities. Results are representative of 3 independent experiments performed in triplicate. Bars represent standard errors.
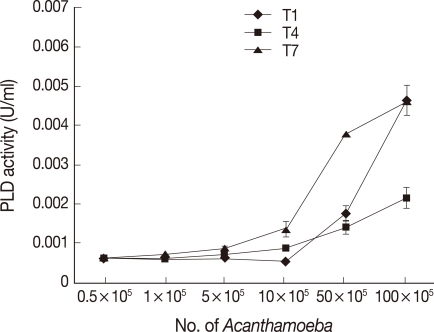
Fig. 3
Acanthamoeba exhibiting optimal phospholipase activities at 37℃. To determine optimal temperatures of Acanthamoeba phospholipases, assays were performed at various temperatures using Acanthamoeba lysates (5×106 amoebae). Note that all Acanthamoeba isolates tested exhibit optimal PLA2 (A) and PLD (B) activities at 37℃ indicating their physiological relevance. Results are representative of 3 independent experiments performed in triplicate. Bars represent standard errors.
Fig. 4
Acanthamoeba exhibiting optimal phospholipase activities at neutral pH. To determine optimal pH of Acanthamoeba phospholipases, assays were performed at various pH using Acanthamoeba lysates (5×106 amoebae). Note that all Acanthamoeba isolates tested exhibit optimal PLA2 (A) and PLD (B) activities at neutral pH indicating their physiological relevance. Results are representative of 3 independent experiments performed in triplicate. Bars represent standard errors.

Fig. 5Phospholipase inhibitors blocking the environmental isolate-mediated binding to HBMEC but not the clinical isolates-mediated binding. The results revealed that both phospholipase inhibitors restricted environmental isolate (C)-mediated binding to HBMEC. However, neither inhibitors had any effects on the clinical isolates (A, T1; B, T4). Results are representative of 3 independent experiments performed in duplicates. Bars represent standard errors.
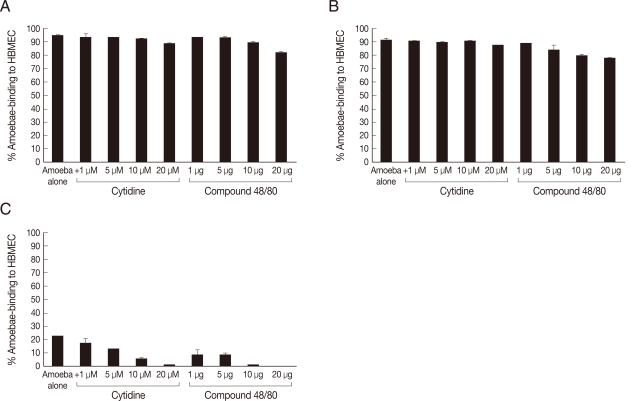
Fig. 6Compound 48/80 partially inhibiting encephalitis isolate-mediated HBMEC cytotoxicity. The results revealed that cytidine 5'-diphosphocholine had no significant effect on Acanthamoeba-mediated HBMEC cytotoxicity. Similarly, compound 48/80 had no effect on Acanthamoeba keratitis isolate (B) but partially inhibited encephalitis isolate (A)-mediated HBMEC cytotoxicity. C is a environmental isolate of T7. Results are representative of 3 independent experiments performed in duplicates. Bars represent standard errors.