Electron Microscopy of the Separated Outer Tegument of the Sparganum and Its Antigenicity
Article information
Abstract
The author reported previously on separation of the outer tegument of the spargana (plerocercoids of Spirometra mansoni) using high concentration of urea solution. To determine which layer of the tegument is separated by this method, an electron microscopic analysis has been processed in this study. It was confirmed that the basement layer of the tegument is separated from the parenchyme of the sparganum. In addition, the antigenicity of the separated outer tegument against the human sparganosis patient sera was evaluated. Numerous antigenic proteins, including 16 and 55 kDa proteins, were noticed in the separated tegument; however, there were no diagnostic 31/36 kDa molecules in this tegument. The molecules reactive with the patient sera in the tegument are to be characterized in future studies.
After infection in the second intermediate host, the spargana (plerocercoids of Spirometra mansoni) worms penetrate the intestinal wall and migrate through the peritoneal cavity within a few hours. Finally, worms reside at various localities, including the subcutaneous tissues, and form a parasitic nodule [1]. The author published on the separation of the outer syncytial layer from the parenchyma of the sparganum using urea solution [2], and the ultrastructure of the cestode tegument using transmission electron microscope (TEM) has been well known. The present study, using TEM, was performed to determine which layer is separated from the parenchyma of the sparganum by urea solution. The results may be important for future functional studies of the cestode tegument, including the physiological nutrient transport and antigenicity against the human sera.
The 31/36 kDa molecules are well known antigens for the diagnosis of sparganosis [3] which are immunolocalized in syncytial and tegumental cells of the worm [4]. Therefore, in consideration of the parasite's tegumental antigenicity, we performed immunoblot to confirm whether the 31/36 kDa molecules are located in the separated tegument and which molecules in the tegument-extracted samples are commonly reactive against the sparganosis sera.
Spargana were collected in the viscera, abdominal cavity, and muscles of naturally infected grass snakes caught in Korea. After washing with sterile physiologic saline, the worms were stored at 4℃ until infection. Spargana from snakes were used to infect BALB/c mice to maintain the worm's good viability. After 1 month, the infected mice were sacrificed, and the spargana were collected from the subcutaneous tissues. Worms were individually added to a culture dish containing RPMI medium supplemented with 3 M urea, 1 mM EDTA, and 30% (v/v) fetal bovine serum [2]. After incubation at 37℃ for 3 hr, worms were washed with PBS (0.1 M, pH 7.4) and fixed in 2.5% glutaraldehyde for 2 hr followed by 1% osmium tetroxide (OsO4) for 1 hr. After rinsing with PBS, the samples were dehydrated using increasing concentrations of ethanol and propylene-oxide. The tegument of the worm was then embedded in synthetic resin (Epon 812). Thin and ultrathin sections were prepared with a Reichert-Jung ultramicrotome (Leica Microsystems, Heidelberg, Germany). Cross-sections of 1 µm in thickness were stained with 1% toluidine blue, and ultrathin sections of 60-70 nm in thickness were stained with uranyl acetate and basic lead citrate. The samples were observed with a H-7650 electron microscope (Hitachi, Tokyo, Japan).
After preparation of the tegumental proteins using the protein extraction buffer (Proprep, Intron Biotechnology, Seoul, Korea), total protein concentrations were measured using the BCA Protein Assay (Intron Biotechnology). Immunoblot was done by conventional methods. Briefly, the tegumental proteins were separated by 7.5-15% SDS-PAGE and then transferred onto polyvinylidene difluoride (PVDF; Millipore, Bedford, Massachusetts, USA) membranes. Membrane strips were then reacted with sparganosis patient sera and other parasite-infected patients sera at a 1:100 dilution in casein buffer (0.5% casein, 20 mM Tris, 150 mM NaCl) and washed with PBS/T (pH 7.4, 0.5% Tween-20). The membranes were then reacted with peroxidase-conjugated anti-human IgG antibody (Fc specific; Cappel, Westchester, Pennsylvania, USA) diluted 1:2,000 in casein buffer. Finally, colors were developed with 4-chloro-1-naphthol (Sigma, St. Louis, Missouri, USA). All reagents used in this study were purchased from Sigma.
As shown in Fig. 1A, the toluidine blue-stained sparganum was clearly seen by light microscopy and also the tegumental separation was noticed almost around the sparganum. The rectangular area was processed for TEM analysis. As shown in Fig. 1B, non-treated control, the tegument of the sparganum was seen, and there was normal architecture of spear-shaped microtriches (mt) which were consistent with the results of the anterior portion of the worm [5], basal layer (BL), and muscle layer (ML). After the tegumental separation using urea solution, microtriches seemed to maintain their normal structures (Fig. 1C), and the basal layer was separated clearly (* in Fig. 1D, E). The sparganum had severely wrinkled folds on the tegument, and some folding portions seemed to be still attached to the parenchyme of the sparganum (Fig. 1A). However, by electron microscopy, it has been confirmed that the separation of the sparganum tegument by urea was clearly complete (Fig. 1D, E).
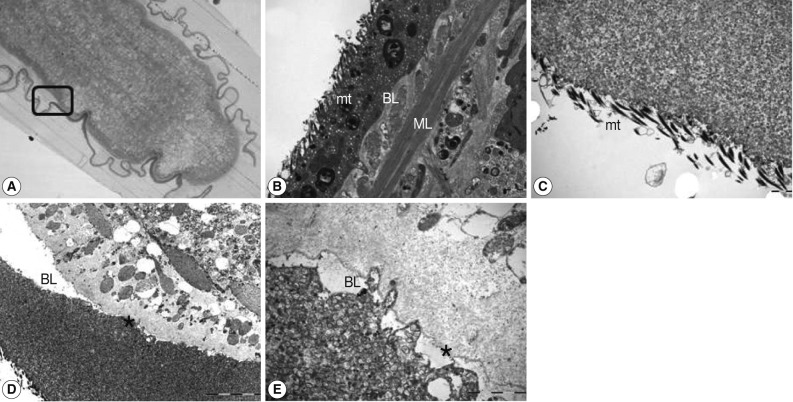
Whole sectional view of the tegument of a sparganum worm. The tegument consisted of a plasma membrane, including the spear-shaped microtriches (mt), protoplasmic layer, basement layer (BL), and muscle layer (ML). The rectangular area in (A) was processed for electron microscopic analysis. Note the asterisk (*) which means the separation of the basement layer (D) from the worm parenchyme (E). Magnification at ×40 (A), ×10,000 (B), ×12,000 (C), ×4,000 (D), and ×20,000 (E). Scale bars: 2 µm in (B), (C), and (E), 10 µm in (D).
Western blot was processed on the proteins of the sparganum as shown in Fig. 2. Numerous reactive protein bands against human sparganosis sera were noticed in the crude extracts of the sparganum (Fig. 2B), including 31/36 kDa molecules which are known as highly antigenic molecules in sparganosis [3]. However, there were no 31/36 kDa molecules in the tegument-extracted sample (Fig. 2A). This means that 31/36 kDa antigenic molecules may be secreted molecules from the parenchymal cells but not present in the tegumental cells. This finding was not consistent with the previous report [4] in which 31/36 kDa molecules were immunolocalized at the syncytial tegument and tegumental cells. In addition, no commonly reactive molecules were noticed in the tegument-extracted samples against sparganosis sera, and possibly 16 kDa and 55 kDa molecules in the separated tegument are candidates for useful antigens in the diagnosis of sparganosis. These results suggested that various immune responses against tegumental molecules of the sparganum occur in the early phases of infection, and more highly specific antigenic molecules, such as 31/36 kDa proteins, are secreted later from the parenchyme of the sparganum.
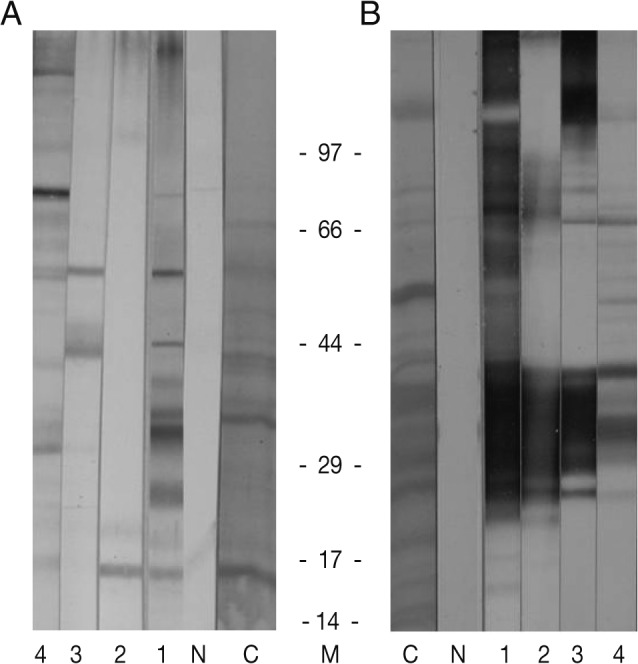
Immunoblot findings of the tegument (A) and crude extracts (B) of the sparganum against human infected sera. Note that several highly reactive antigenic molecules in the tegument proteins (A) and 31/36 kDa proteins in crude extracts (B). C, crude extracts; N, normal sera; 1-4, sparganosis patient sera.
The characteristic structure of the tapeworm tegument is known to participate in the absorption of nutrients, excretion of waste metabolites, protection from host digestive enzymes [6,7], and immunolocalization of antigenic molecules [4]. Also, in electron microscopy, the tegument of the sparganum is covered by numerous microtriches and glandular cells which may be used for migration of the worms in the host [2,8], and the tegument will represent an initial antigenic contact with the host. In this study, the presence of potent antigenic molecules in the tegument of the sparganum has been confirmed by immunoblot using human sparganosis sera. Further characterization of these antigenic molecules should be performed.
ACKNOWLEDGMENT
The author thanks Mr. Je-Young Ryu for his technical assistance.