Tegumental Ultrastructure of Adult Gynaecotyla squatarolae (Digenea: Microphallidae)
Article information
Abstract
Gynaecotyla squatarolae (Digenea: Microphallidae) adult flukes were recovered from experimental chicks at day 4-6 post-infection and their tegumental ultrastructure was observed with a scanning electron microscopy. They were pyriform in shape, and their anterior halves were concaved ventrally. The whole body surface was covered with tegumental spines, which were wide and 16-17 digitated between oral and ventral suckers. The density of spines and number of digits decreased posteriorly. The oral sucker was subterminal and the excretory pore was at the posterior end of the worm. Two ventral suckers were similar in appearance and protruded near midline of the worm. The genital atrium was dextral to the small ventral sucker. The dorsal surface was covered with tegumental spines, but the spines were sparser than on the ventral surface. On the middle portion of the dorsal surface, a small opening presumed to be the Laurer's canal was seen. From these findings, it has been confirmed that the adult G. squatarolae has unique characteristics in the surface ultrastructure.
INTRODUCTION
Gynaecotyla squatarolae (Digenea: Microphallidae) was originally discovered in the small intestines of birds [1], and the second intermediate host was shown to be the shore crab, Macrophthalmus japonicus and Macrophathalmus dilatatus in Japan [2]. In the Republic of Korea, the metacercariae of G. squatarolae were discovered from M. dilatatus [3], and the ruddy turnstone, Arenaria interpres interpres, was reported as a natural definitive host [4]. Whereas the majority of trematodes require a definitive host to develop into adults, microphallid metacercariae have been shown to undergo extensive organogenesis. For example, Microphallus opacus, a member of Microphallidae, is able to complete its life cycle without the involvement of a vertebrate host [5]. The metacercariae of G. squatarolae excyst quickly after extraction from the crab, and possess a primitive ovary and testes [3]. The first intermediate host of Gynaecotyla sp. is yet unknown, and thus efforts to elucidate the biology of G. squatarolae should be exerted.
The surface ultrastructure of trematodes provides us with insight into their morphological and taxonomical characteristics, and it has been demonstrated that the shape and distribution of tegumental spines differ by species [6]. Gynaecotyla sp. belongs to the family Microphallidae, in which more than 17 genera have been known [3], and had unique morphological characteristics, including 2 ventral suckers, separate male and female genital pores, and a well-developed genital atrium. However, only a few studies have been conducted regarding the tegumental ultrastructure of microphallid trematodes. The metacercariae of Microphallus abortivus were observed via transmission electron microscopy (TEM) and scanning electron microscopy (SEM) [7], and the structures of the metacercarial cyst of 4 microphallid trematodes have been previously reported [8]. Pina et al. [9] reared the larval stage of Cercaria sevillana into metacercariae in the crustacean, and concluded that it was a microphallid fluke, particularly Gynaecotyla longiintestinata, with the aid of SEM [9]. However, no information is currently available regarding the surface ultrastructure of adults of Gynaecotyla sp. Therefore, the present study was undertaken to observe the tegumental ultrastructure of G. squatarolae adults.
MATERIALS AND METHODS
The metacercariae of G. squatarolae were isolated from brackish water crabs, Macrophthalmus dilatatus, purchased at a local market in Taean-eup, Chungcheongnam-do, the Republic of Korea, February 2008. Total 500 metacercariae were orally fed to 10 chicks using a gavage needle, and the chicks were sacrificed on days 4 and 6 post-infection (PI). The small intestines were longitudinally opened and loaded on Baermann's apparatus. Adult worms were recovered and washed several times with physiological saline and fixed in 2.5% glutaraldehyde at 4 ℃. The specimens were dehydrated in a graded ethanol series, dried in a critical point dryer (ABT, Hitach HCP2, Hitach Co., Tokyo, Japan), and mounted on stubs. The dried flukes were coated with gold using an ion sputtering coater (Hitach 2-1030, Hitach Co.) and observed with a scanning electron microscope (HITACH 54705, Hitach Co.) at an accelerating voltage of 5.0 kV.
RESULTS
The adult G. squatarolae was pyriform in shape and concaved ventrally. The ventral concavity was more prominent in anterior half of the body. The oral sucker was at the anterior subterminal end. Two ventral suckers were located at a mid-median level. The posterior half of the body was wrinkled with longitudinal folds, and the excretory pore was opened at the posterior terminal end (Fig. 1A). The whole body surface was beset densely with scale-like tegumental spines, and some sensory papillae were distributed among them. On the ventral surface, the oral sucker was elliptical, 50 × 55 µm in size (Fig. 1B). Three to 4 rows of tegumental spines having 6-9 digits were distributed on the lateral margin of the oral sucker. The region anterior to the oral sucker was devoid of tegumental spines, on which sensory papillae consisting of type I (ciliated, knob-like) and type II (aciliated round swelling) were found (Fig. 1B). The tegumental spines just posterior to the oral sucker were wide, with 16-17 digitated projections (Fig. 1C). On the tegument between the oral and ventral suckers, the spines were similar to those of the oral sucker in appearance, but the density of spines was slightly decreased. In addition, the number of digits decreased to 13-14.
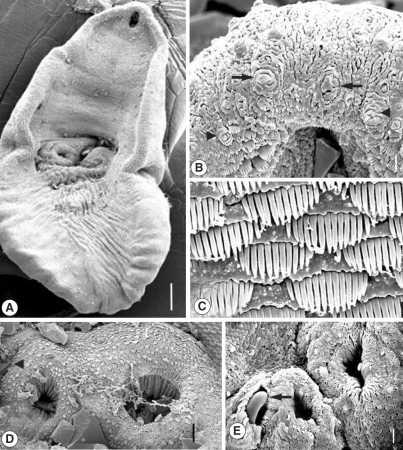
Scanning electron micrographs of Gynaecotyla squatarolae (ventral view). (A) A whole adult worm. Oral sucker and two ventral suckers are seen. Bar = 83.3 µm. (B) Tegument around the oral sucker. Note the type I sensory papillae (arrowheads) and aciliated type II sensory papilla (arrows). Bar = 5.0 µm. (C) Tegument on the posterior of oral sucker, showing dense distribution of tegumental spines, having 16-17 digits. Bar = 1.7 µm. (D) Two ventral suckers are seen. Large ventral sucker (arrow) on the left, and small one on the right. Right to small ventral sucker located the genital pore (arrowhead). Inside the suckers, numerous wrinklings formed the column-like structures. Bar = 1.7 µm. (E) Another worm showed the genital atrium dextral to the small ventral sucker. A curved cirrus (arrow) is observed inside it. Bar = 9.5 µm.
The small ventral sucker was on the right and the larger one on the left side (Fig. 1D). They were spherical in shape, and numerous spines having 3-6 digits surrounded their lips. They had several sensory papillae on the lips. Inside the 2 ventral suckers, numerous wrinkles were observed, resembling regularly distributed columns. The genital atrium was dextral to the small ventral sucker, and the curved cirrus protruded from it (Fig. 1E). The spines on the posterior body surface were sparsely distributed and fork-shaped with 3-5 digits.
The dorsal surface was covered densely with tegumental spines (Fig. 2A). On the middle portion of the dorsal surface, a small opening was observed. It was a curved furrow, lying transversely, and supposed to be the Laurer's canal (Fig. 2B). On the anterior portion of the body, the spines were shorter than those of the ventral surface, and had 12-14 digits (Fig. 2C). The number of digits decreased to 6-7 on the middle portion (Fig. 2D) and finally became one digit per spine at the terminal portion (Fig. 2E).

Scanning electron micrographs of Gynaecotyla squatarolae (dorsal view). (A) Dorsal surface of the body. The Laurer's canal like opening is seen (arrow). Bar = 83.3 µm. (B) Magnification of an arrow mark in (A). It looked like a deep furrow, lying transversely. Bar = 1.7 µm. (C) Tegument on the dorsal surface near the anterior end, showing distribution of tegumental spines, having 12-14 digits. Bar = 1.7 µm. (D) Tegument on the dorsal surface near the middle portion of the worm. The number of digits was in the range of 6-7. Bar = 1.7 µm. (E) Tegument of dorsal surface, near the posterior end of body. Each spine has a digit, and the density of spines was very low. Bar = 1.7 µm.
DISCUSSION
A characteristic feature of G. squatarolae is possessing 2 ventral suckers; the smaller right one and the large left one. The findings in the present specimen were consistent with the notion that the left acetabulum is generally larger than the right one in adult Gynaecotyla adunca, and in the metacercariae of G. longiintestinata [9,10]. While the genital sucker of Heterophyes nocens was the modification of the genital atrium [6], an investigation on G. squatarolae showed that the 2 suckers were similar in appearance to each other and that numerous wrinkles radiated deep inside. However, this finding did not mean that the 2 suckers were equally involved in attachment to the host tissue, and it should be proved later.
In the present study, the tegument between the oral and ventral suckers was covered with wide spines possessing 16-17 digits. The number of digits was similar to that of Pygidiopsis summa (14-16) [11], but they differed in morphology. While the spines of P. summa were broom brush-shaped [11], those of G. squatarolae were wide and rectangular shaped, and each digit looked to be separated from each other. These wide spines were also observed in metacercariae of G. longiintestinata [9], thereby suggesting the taxonomic significance. The division of tegumental spines has been associated with the abrasion of the host tissue and locomotion in the host [12]. The spines of Heterophyopsis continua were surmised to perform a function of migratory behaviors [13], but in Echinostoma caproni the multipointed spines started to develop at about the time when the body growth was less rapid [14]. Considering that adults of E. caproni do not leave the mouse small intestine, the development of multipointed spines was probably not associated with a change in habitat. On the contrary, the excysted metacercariae of Gynaecotyla had already developed tegumental spines similar to those of G. squatarolae adults [9], thereby suggesting a relationship with locomotion. However, the significance of this phenomenon remains unclear. The wrinkles located at the posterior half were probably formed by the vitellaria or the uterus having intrauterine eggs.
On the dorsal surface of G. squatarolae, we detected a small opening near the middle portion and suggested it to be the Laurer's canal. However, by means of light microscopy no Laurer's canal was detected in G. squatarolae [1,3]. It was also not observed in G. adunca [10]. This may reflect limitations of light microscopy. However, the presumed Laurer's canal of our specimen was different from that of Metagonimus miyatai, which was surrounded by several layers of sphincter-like structures, and numerous sperms were entering through it [15]. Since no sperm was observed near the opening of the present specimen, it is suspected to be a vestigial Laurer's canal resulting from an absence of cross-fertilization in G. squatarolae. The presence of the Laurer's canal in G. squatarolae requires further studies.
Conclusively, the surface ultrastructure of G. squatarolae evidenced some unique characteristics, such as wide spines with numerous digits on the ventral surface, and more research on this species should be clearly conducted.