Molecular Detection of Toxoplasma gondii in Haemaphysalis Ticks in Korea
Article information
Abstract
Toxoplasma gondii are intracellular protozoa that can cause neurological disease or death in fetuses and even in immunocompromised human adults. Ticks are recognized as vectors of many microorganisms including viruses, bacteria, and protozoa. Recent studies detected T. gondii in various tick species in many countries. In this study, we performed PCR detection of the T. gondii B1 gene from Haemaphysalis ticks collected from vegetation in 4 localities, Wonju, Gunsan, Miryang, and Yangsan, in Korea. We analyzed DNA from 314 ticks (268 Haemaphysalis longicornis and 46 Haemaphysalis flava) and the B1 gene of T. gondii was detected in 13 of these. The detection of T. gondii in ticks differed significantly by region (P=0.021). T. gondii was detected in the following percentages of collected ticks: 3.7% (7 of 189) in Gunsan, 10% (5 of 50) in Wonju, 16.7% (1 of 6) in Yangsan, and 0% (0 of 69) in Miryang. The detection of T. gondii in ticks was not associated with tick species or development stage. This is the first report of T. gondii detection in ticks in Korea. Our results provide important information necessary to understand toxoplasmosis transmission.
Toxoplasma gondii can cause severe neurological disease or death in developing human fetuses and in immunosuppressed patients; however, infected immunocompetent individuals are usually asymptomatic [1]. The seroprevalence of T. gondii in Korean people was reported as 5.6% in 1960, 7.2% in 1983, 7.7% in 1999, 6.6% in 2000, 6.7% in 2009, and 8.6% in 2016 [2–7].
T. gondii has been detected in Korean wild animals such as feral cats, Chinese water deer, roe deer, and raccoon dogs [8–10], and in domesticated animals such as cattle, horses, rabbits, and dogs in Korea [11–14]. These animals include herbivores, which raises the possibility that bloodsucking arthropods such as ticks could transfer T. gondii [15]. Ticks are vectors of many microorganisms including viruses, bacteria, and protozoa.
Recently, T. gondii has been detected in various tick species [15], such as Dermacentor reticulatus and Ixodes ricinus in Poland [16–23], Amblyomma in the Republic of Chad [24], and Haemaphysalis longicornis in China [25]. H. longicornis is the dominant tick species in Korea. In addition, many experiments have demonstrated the possibility of toxoplasmosis transmission via ticks [25–30].
In this study, we detected T. gondii DNA in Haemaphysalis ticks collected from vegetation in 4 localities in Korea. Ixodid tick H. longicornis is endemic to China, Korea, Japan, Russia, Australia, and New Zealand and is an important vector of protozoal parasites including Babesia ovata, Babesia gibsoni, Theileria sergenti, and Theileria mutans [31,32]. However, T. gondii has not been found in Korean ticks to date.
Ticks were collected from the vegetation by flagging. The tick collection areas were selected based on topographical, temperature, seasonal and weather aspects, among others. Ticks were collected from August 2014 to October 2016 in various Korean provinces to investigate possible regional characteristics: Wonju (Gangwon-do Province; 37.389545, 127.801770), Gunsan (Jeollabuk-do Province; 36.006237, 126.807751), Miryang (Gyeongsangnam-do Province; 35.475082, 128.780564), and Yangsan (Gyeongsangnam-do Province; 35.286111, 129.027625) (Fig. 1; Table 1). Species identification of collected ticks was performed by examination under a dissecting microscope according to Yamaguti et al. [33], and confirmed by PCR amplification of the tick DNA and sequencing of the 5.8S rRNA internal transcribed spacer 2 region using the primers HITS2-F (5′-GGTGCTCGAGACTCGTTTTG-3′) and HITS2-R (5′-ATTCGCGGTTTACGAGAGAA-3′) [34]. DNA was extracted from each collected tick using a NucleoSpin DNA Insect kit (Macherey-Nagel, Düren, Germany) according to the manufacturer’s instructions, and stored at −20˚C until use. To detect the T. gondii B1 gene in ticks, nested PCR was performed with 2 primer pairs: S1 (5′-TGTTCTGTCCTATCGCAAC G-3′) and AS1 (5′-ACGGATGCAGTTCCTTTCTG-3′), which amplify a 580-bp fragment; and S2 (5′-TCTTCCCAGACGTGGATTTC-3′) and AS2 (5′-CTCGACAATACGCTGCTTGA-3′), which amplify a 530-bp fragment [35]. The PCR products were sequenced (Bionics Co., Seoul, Korea). A BLAST search was used to compare the obtained sequences to those available in GenBank (USA).
Four localities of tick collection. Ticks were collected from the vegetation by flagging in Wonju, Gunsan, Miryang, and Yangsan in Korea.

Summary of Haemaphysalis ticks collected from vegetation in four localities in Korea
A total of 314 ticks from 4 regions were analyzed in this study, comprising 268 H. longicornis and 46 H. flava (Table 1). For 13 of these 314 ticks, agarose gel electrophoresis of PCR products revealed a band pattern at 530 bp (Fig. 2), which indicated the presence of the T. gondii B1 gene. The DNA sequence of these PCR products showed greater than 99.7% identity with the published B1 gene sequence of T. gondii (MH744807.1) (Table 2).
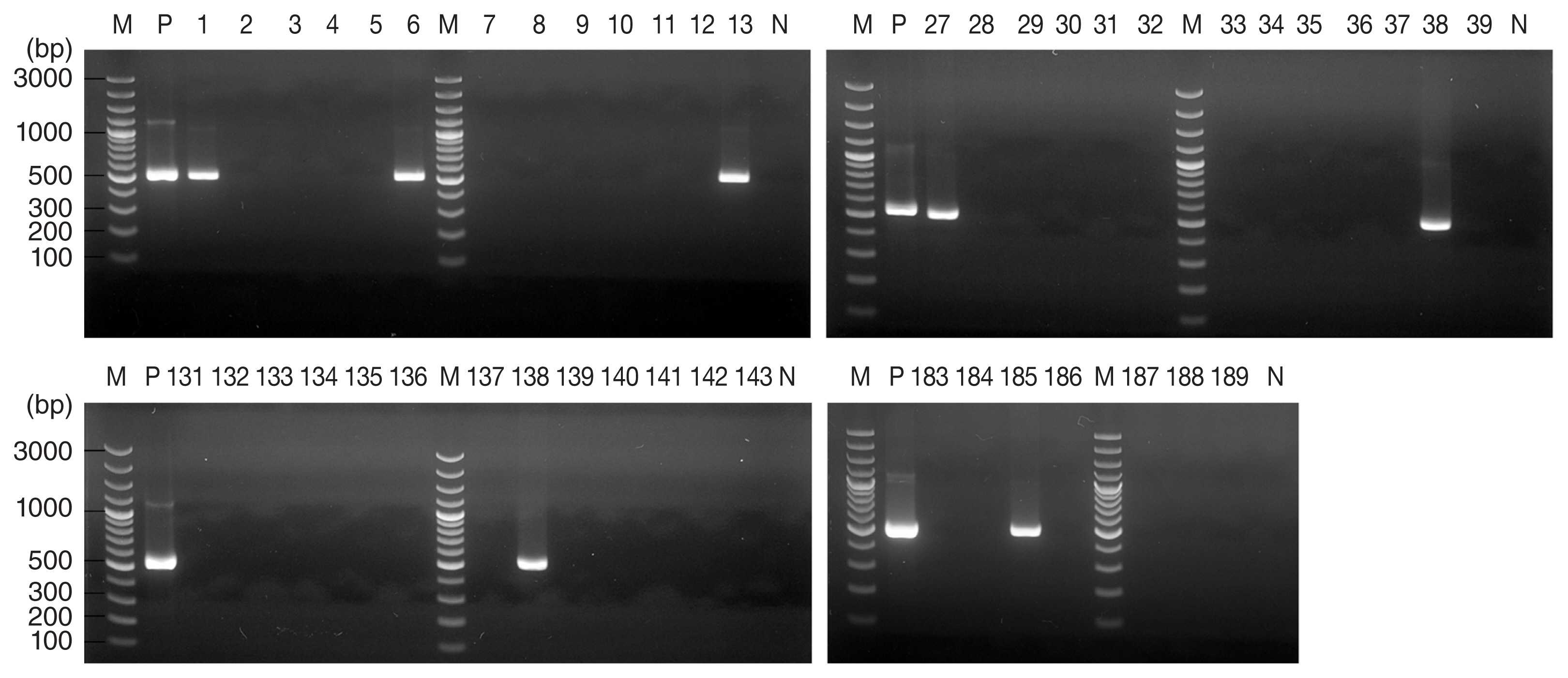
Agarose gel electrophoresis results of Toxoplasma gondii B1 gene PCR products from Haemaphysalis ticks collected from Gunsan, Korea. PCR products from 7 of 189 tick DNA samples indicated the presence of T. gondii. M, marker; P, positive control (T. gondii DNA); N, negative control.

Details of Toxoplasma gondii-positive Haemaphysalis ticks
The detection of T. gondii in ticks differed significantly by collection locality (P=0.021) (Table 3). The T. gondii B1 gene was detected in 7 (3.7%) of 189 ticks collected from Gunsan, in 5 (10%) of 50 ticks collected from Wonju, and in one (16.7%) of 6 ticks collected from Yangsan. T. gondii was not detected in the 69 ticks collected from Miryang.

Association of variables related to Toxoplasma gondii-positive of ticks
The detection rate of T. gondii was 4.1% (11 of 268) in H. longicornis and 4.3% (2 of 46) in H. flava. This indicated there was no association between the species of tick and the detection of T. gondii (Table 3). The detection rates of T. gondii in tick adults, nymphs, and larvae were 14% (2 of 14), 4.9% (5 of 103), and 3.0% (6 of 197), respectively. However, the detection rate of T. gondii among these 3 development stages was not statistically different (P=0.113).
In this study, the detection rates of T. gondii in H. longicornis and H. flava were lower than the previously reported detection rate of T. gondii in H. longicornis (9.2%; 39 of 422) from Qingdao, Shandong Province, China [25]. This study did not find T. gondii in Haemaphysalis larvae, but our results showed the presence of T. gondii in 3% (6 of 197) of larvae. Previous studies have reported the presence of T. gondii in many tick species. In north-western Poland, 12.7% of 259 Ixodes ricinus were infected with T. gondii [16]. The same study demonstrated that inoculation with homogenate of ticks caused toxoplasmosis in 44 of 60 mice [16]. T. gondii DNA was detected in 2.8% of adult I. ricinus in eastern Poland [17], and in 64.9% of I. ricinus adults and nymphs in Upper Silesia, Poland [18]. T. gondii DNA was also found in 3% (52 of 1,737) of I. ricinus feeding on ponies and in 10.2% (38 of 371) of I. ricinus from vegetation including 7 larvae [19]. T. gondii DNA was detected in 3.2% (21 of 664) of adult Dermacentor reticulatus, a human-biting tick, collected from vegetation [20].
Several laboratory experiments have demonstrated the transmission of toxoplasmosis by ticks. Amblyomma americanum, Dermacentor variabilis, and Dermacentor andersoni transmitted T. gondii by blood feeding in the laboratory animals [26]. D. andersoni showed transovarial transmission of T. gondii to offspring [26]. I. ricinus can transmit T. gondii to mice through blood feeding [27]. T. gondii was reported to survive in the body of H. longicornis for more than 10 days [25,28] and T. gondii was transmitted to the host by ingestion, but not by blood-feeding, of infected ticks [25].
T. gondii has previously been detected in wild animals and livestock in Korea. T. gondii oocysts were found in 0.89% (5 of 563) of feral cat feces [8]. T. gondii IgG test results were positive in 12% (6 of 50) feral cats [9]. A PCR assay indicated the presence of T. gondii in 47.2% (50 of 106) of feral cats and 46.3% (64 of 138) of dogs [14]. The incidence of T. gondii seropositivity was 10.8% in Chinese water deer, 4.3% in raccoon dogs, 16.7% in roe deer [10]. Furthermore, the T. gondii B1 gene was detected in the blood of 16.2% (23 of 142) of rabbits from breeding farms in Korea [11]. The incidence of T. gondii seropositivity was 0.5% (3 of 568) in cattle [12], and 2.9% (24 of 816) and 2.6% (5 of 191) in 2 studies on horses [13,36].
It is well known that T. gondii infection can be caused by contact with T. gondii-oocyst-shedding cats and ingestion of T. gondii-contaminated food [37]. However, we suggest the possibility that the tick is a potential vector for transmission of T. gondii, based on the worldwide presence of T. gondii in ticks, experimental evidence of toxoplasmosis transmission by ticks, and cases of T. gondii infection in several herbivorous wild animals and livestock [15].
This is the first report of T. gondii detection in ticks in Korea. T. gondii was found in all lifecycle stages of both H. longicornis and H. flava ticks collected from vegetation in Gunsan, Wonju, and Yangsan. This study provides important information necessary to understand toxoplasmosis transmission. We suggested a new transmission cycle of T. gondii as vector-borne in wild and domestic animals.
Supplementary Information
DNA sequences of B1 gene of Toxoplasma gondii detected in ticks
ACKNOWLEDGMENTS
This study was supported by a National Research Foundation of Korea (NRF) grant funded by the Korean Government (Ministry of Education, Science and Technology; grant no. 2019R1A2B5B01069843).
Notes
CONFLICT OF INTEREST
The authors have no conflicts of interest to declare.