Trichinella Infection Ameliorated Vincristine-Induced Neuroinflammation in Mice
Article information

Abstract
Vincristine (VCR) is a chemotherapeutic agent widely used in treatment of malignancies. However, VCR has a limitation in use since it commonly causes a painful neuropathy (VCR-induced peripheral neuropathy, VIPN). Inflammatory cytokines secreted by immune cells such as macrophages can exacerbate allodynia and hyperalgesia, because inhibiting the inflammatory response is a treatment target for VIPN. In this study, we investigated whether Trichinella spiralis, a widely studied helminth for its immunomodulatory abilities, can alleviate VCR-induced allodynia. Von Frey test showed that T. spiralis infection improved mechanical allodynia at 10 days after VCR injection. We further observed whether the difference was due to mitigated axon degeneration, but no significant difference between the groups in axonal degeneration in sciatic nerves and intra-epidermal nerve fibers was found. Conversely, we observed that number of infiltrated macrophages was decreased in the sciatic nerves of the T. spiralis infected mice. Moreover, treatment of T. spiralis excretory-secretory products caused peritoneal macrophages to secrete decreased level of IL-1β. This study suggests that T. spiralis can relieve VCR-induced mechanical allodynia by suppressing neuroinflammation and that application of controllable degree of helminth may prove beneficial for VIPN treatment.
INTRODUCTION
Vincristine (VCR) is a vinca alkaloid, which is a chemotherapeutic agent widely used in treatment of several kinds of cancer including hematological malignancies. However, it has neurotoxicity as its common side effect. Painful VCR-induced peripheral neuropathy (VIPN) causes serious adverse effects not only in patient’s quality of life but also in cancer treatment [1,2]. There are still few effective drugs that can prevent or improve VIPN.
Although the mechanisms how VCR induces neurogenic pain have not fully been elucidated, complex mechanisms appear to be involved in this process. Since VCR is a microtubule depolymerizing agent, it can directly induce disturbance of axonal transport, possibly triggering sterile alpha and TIR motif containing 1 (Sarm1)-dependent axonal degeneration [3]. VCR treatment induces an increase in monocytes/macrophages in nerves, and these recruited cells contribute to exacerbate VCR-induced allodynia [4].
Macrophages secret inflammatory cytokines including interleukin (IL)-6, IL-1β, and tumor necrosis factor (TNF)-α. Since these cytokines increase pain sensitivity, drugs that inhibit neuroinflammation are considered potential therapeutic candidates.
Parasitic helminth is harmful to the host, but the versatile nature of helminth can be used in ways that are beneficial to humans. Recently, therapeutic availability of helminths is drawing attention for inflammatory disorders which are difficult to treat [5,6]. Parasitic helminths have evolved to have ability to escape from host’s immune system. A lot of helminths has a powerful ability to modulate host immune system [7–9]. Trichinella spiralis, for example, is one of the most commonly studied helminths in animal model for autoimmune diseases [10–12]. T. spiralis infection causes host to get resistance to autoimmune diseases. T. spiralis infection inhibits inflammatory activation of immune cells in several manners. Protective effect for experimental autoimmune encephalitis appears to be related to production of high level IL-10 by increasing Foxp3+ regulatory T cells [13]. Excretory-secretory products (ESP) of T. spiralis can also prevent secretion of pro-inflammatory cytokines including IL-1β in macrophages and suppress dextran sodium sulfate-induced colitis of mice [14,15].
Although T. spiralis infection improves CNS inflammation [13,16], little information is reported on the effects of T. spiralis infection on neuroinflammation of peripheral nervous system (PNS). In this study, we explored whether T. spiralis infection can improve VIPN in mice.
MATERIALS AND METHODS
T. spiralis infection
The T. spiralis strain (isolate code ISS623) used in this study has been cultured in Pusan national university [11]. For infection of the parasite, 250 T. spiralis larvae per mouse were orally administered to female 4–6 weeks old C57BL/6 mice. All animal experiments were approved by the Dong-A University Committee on animal research (DIACUC-22-10), and follows the guidelines for animal experiments of the Korean Academy of Medical Sciences.
Preparation of excretory-secretory products (ESP)
T. spiralis ESP were prepared as previously described [17]. Briefly, mice were sacrificed at 4–6 weeks post infection, after which, skinned carcasses were chopped into pieces and digested in 1% pepsin-hydrochloride for 2 h at 37°C with stirring. Larvae were manually collected from digested muscle under a microscopy, after which cleaned in sterile phosphate buffered saline (PBS; 100 mM, pH 7.4) at least 5 times. The larvae were incubated in serum-free RPMI-1640 medium containing penicillin/streptomycin (100 μg/ml) at 37°C for 5 days. We confirmed that all the worms were alive after incubation. After the supernatant was collected, protein concentration is measured using bicinchoninic acid method.
Induction of VIPN
For VIPN induction, vincristine sulfate (Tocris, Bristol, UK) dissolved in physiological saline was injected into peritoneal cavity of T. spiralis infected or uninfected C57BL/6 mice (0.5 mg/kg/day). The treatment was administered for 5 consecutive days followed by a 2 day rest period. The cycle was repeated until the VCR was treated for a total of 18 days [18].
Measurement of mechanical allodynia
Mechanical allodynia was evaluated by von Frey test using ascending stimulus method starting with the 0.16 g filament. The lowest value that causes an avoidance reaction of 50% or more was recorded as paw withdrawal threshold. After the results of both hind paws of each mouse were recorded, the average value was analyzed for group mean values.
Peritoneal macrophages
For harvesting peritoneal macrophage, 10 ml of sterile PBS containing penicillin/streptomycin was injected into peritoneal cavities of C57BL/6 mice. After that, the injected PBS containing peritoneal cells was extracted with a syringe. Then, the cell suspension from the peritoneal cavity was centrifuged at 200×g for 5 min. The pellet was resuspended in RPMI-1640 medium, the number of viable cells was counted, and the cells were seeded into 24-well plate.
Immunofluorescence (IF) staining
After sacrificing the mouse, the sciatic nerve was carefully collected and fixed in 4% paraformaldehyde. After 24 h of fixation, the sciatic nerve was transferred to 20% sucrose for an additional 24 h. Then the sciatic nerve was embedded and frozen in optimal cutting temperature (OCT) compound. The block was sectioned using a cryostat (Leica Biosystems, Nussloch, Germany) at 10 μm thickness. After blocking with PBS containing 0.2% Triton X-100 with 5% bovine serum albumin for 1 h, the slides were incubated with antibodies against CD68 (diluted to 1:3,000, Bio-Rad, Munich, Germany), myelin basic protein (diluted to 1:1,000, Abcam, Cambridge, UK), and PGP9.5 (diluted to 1:1,000, Abcam) for overnight at 4°C. Allexa 488- or Cy3-conjugated secondary antibodies were treated for 2 h at room temperature. For nuclear staining, slides were incubated with 4′, 6-diamidino-2-phenylindole (DAPI) (Sigma, St. Louis, Missouri USA) for 15 min. Zeiss imager M2 with ApoTome II microscope (Carl Zeiss, Oberkochen, Germany) was used to obtain images.
For the analysis of intra-epithermal nerve fiber (IENF), hind paw foot pads were taken from each mouse and immediately immersed in Zamboni’s fixative for 2 h at room temperature. The tissues were washed with PBS 3 times for 10 min and transferred to 20% sucrose. The cryopreserved tissues were sectioned at 50 μm thickness. The number of IENF was counted using 20× objective.
ELISA
Measurement of mouse cytokines in culture media was performed with commercially available ELISA kits. For evaluation of IL-6 and TNF-α, ELISA kits purchased from BD biosciences (Franklin Lakes, New Jersey, USA) were used. IL-1β ELISA kit was from Cusabio (Houston, Texas, USA). All tests were performed in triplicates according to the manufacturer’s instructions.
Western blot analysis
For western blot analysis, sciatic nerves were dissected and lysed for 3 min in RIPA buffer by using TissueLyser LT (Qiagen, Hilden, Germany). The tissue lysates were centrifuged at 9,000×g for 5 min at 4°C and the supernatant was collected. The protein samples (10–30 μg) were separated by 10% reducing SDS-PAGE and transferred onto a nitrocellulose membrane (Amersham Biosciences, Amersham, UK). After blocking with 5% skim milk in Tris-buffered saline with 0.05% Tween-20 (100 mM, pH 7.2) for 1 h at room temperature, the membranes were incubated with antibodies specific to CD68 overnight at 4°C. The membranes were washed 3 times, and then the membranes were incubated with a horseradish peroxidase-conjugated secondary antibody for 1 h at room temperature. Immunoreactive signals were detected using enhanced chemiluminescence solution (GE Healthcare, Chicago, Illinois, USA). The images were obtained using a LuminoGraph 3 imaging system (ATTO, Tokyo, Japan). Quantification of band intensities was performed with CS Analyzer 4 (ATTO).
Statistical analysis
Statistical analysis was performed using GraphPad Prism software (GraphPad software, San Diego, California, USA). P values were from Student’s two-tailed test, and results were given as the mean±SEM. The results with P values less than 0.05 were considered to be statistically significant.
RESULTS
T. spiralis infected mice showed ameliorated VCR-induced mechanical allodynia
To evaluate the mechanical allodynia, von Frey test was performed on 0-, 5-, 10-, and 15-days post VCR injection, respectively. There was no difference between T. spiralis infected (Inf+) and uninfected (Inf-) groups, before VCR injection. No significant difference was also observed until the 5 days post injection. Interestingly, we found that the paw withdrawal threshold at 10 days post injection was significantly higher in the Inf+ group (Fig. 1A). However, in the test results conducted on the 15 days post injection, the statistical significance between groups was no longer observed. This result may suggest that there is little protection against continuous deterioration.
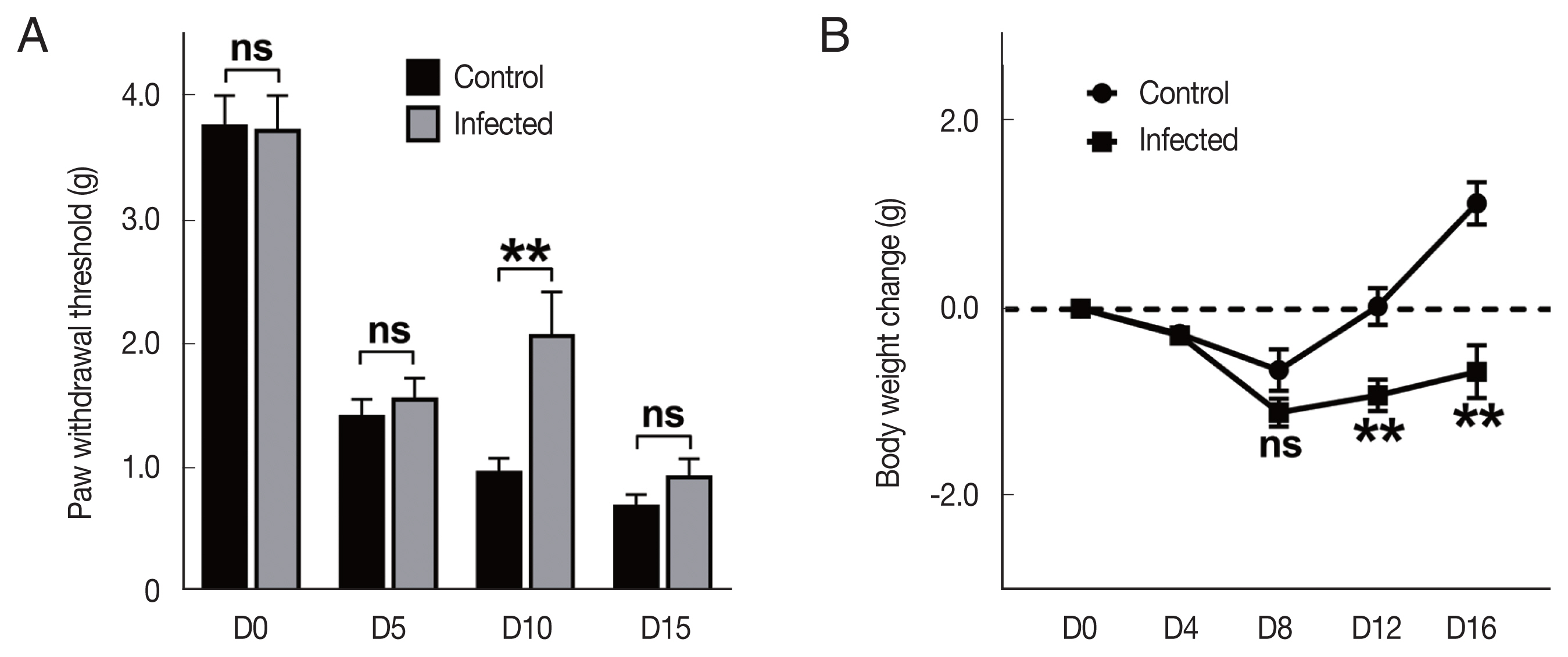
Vincristine treatment time-dependently induced phenotype differences in Trichinella spiralis infected mice. (A) Paw withdrawal threshold score conducted by von Frey test (n=8 each). (B) Changes in body weight after VCR treatment. The values indicate the average weight change in the group compared to the start of each mouse. Control, T. spiralis uninfected VCR-treated group; Infected, T. spiralis infected VCR-treated group. ns, not-significant. **P<0.01 by Student t-test.
We also observed changes in body weight to get information about the general state of the mice. At the beginning of the experiment, body weights of the mice were between 20 and 23 g, and the body weights were measured every 4 days since the VCR injection began. Each group lost their body weight after VCR injection, showing the lowest weight around 8 days, and then slowly regained (Fig. 1B). The time point at which the weight began to recover was similar between the groups, however, the (Inf+) group showed lower weight gain than (Inf-) group. Since the parasitic helminth infection itself can affect mouse weight changes [19,20], this result is probably due to the T. spiralis infection itself rather than to the effect of VCR toxicity.
T. spiralis infection did not affect VRC-induced axonal degeneration
VCR is known to cause axonal degeneration [3], thus we tried to determine whether the difference in mechanical allodynia between the (Inf+) and (Inf-) groups comes from the difference in the degree of axon degeneration.
Sensory nerve conduction study conducted by mice tail showed no significant difference before and after VCR processing in both infected and uninfected groups (Fig. 2A). There was no significant difference in amplitude and conduction velocity of sensory nerve (Fig. 2B, C). These results suggest that no dramatic axonal degeneration in major nerve has occurred that can be reflected in the electrophysical study in this experimental condition.
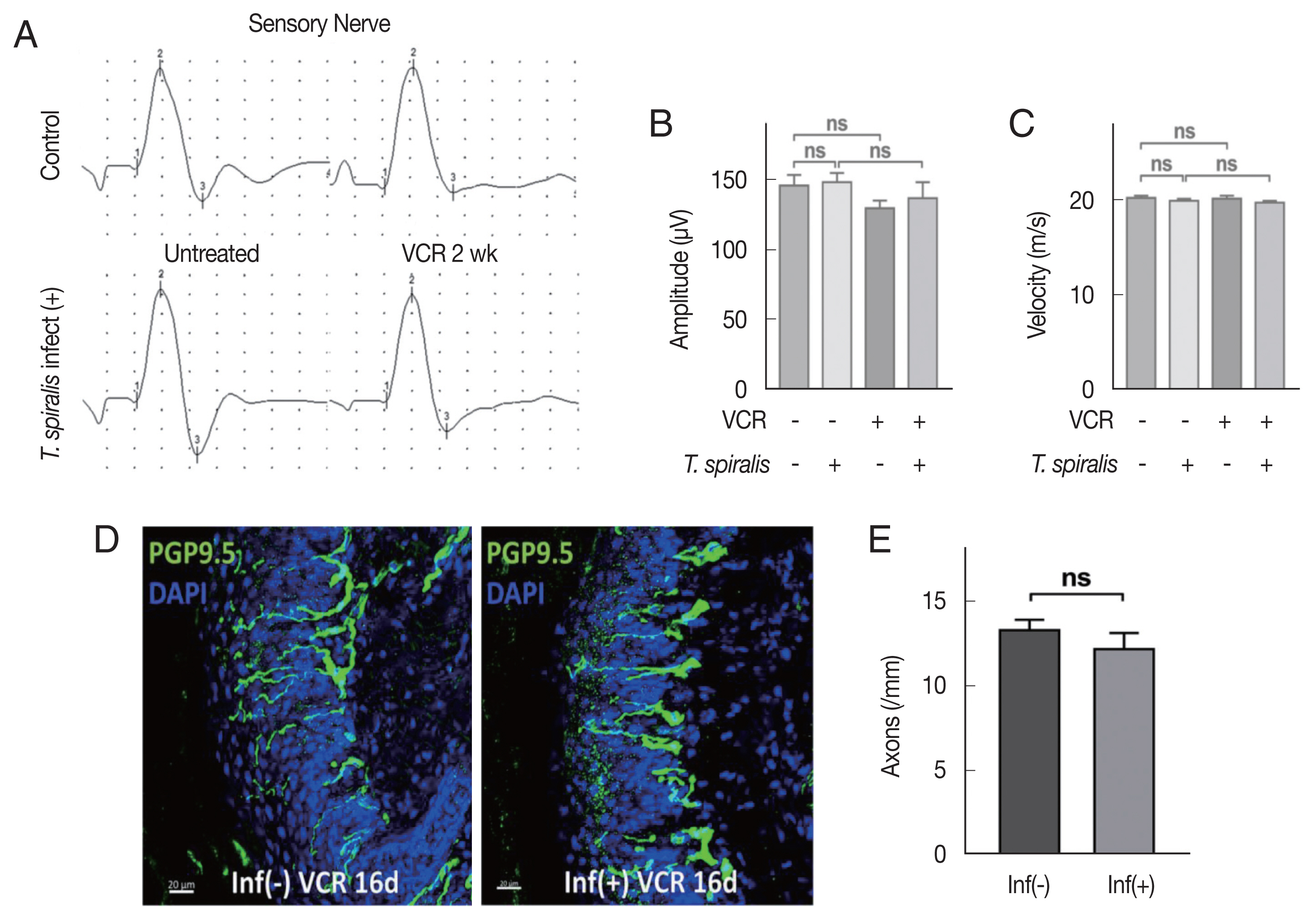
Vincristine treatment did not significantly alter axon degeneration in Trichinella spiralis infected mice. (A) Sensory nerve conduction of caudal nerves. (B) Quantitative analysis of amplitude. (C) Conduction velocity. (D) Representative immunofluorescent images showing intra-epidermal nerve fiber (PGP9.5) in hind paw foot pads of the uninfected (left) and T. spiralis infected (right) mice at 16 days post VCR treatment. (E) Histogram showing quantitative analysis of intra-epidermal axon count. (Inf-); uninfected group, (Inf+); T. spiralis infected group. Scale bar=20 μm. ns, not-significant.
Next, we observed intra-epidermal nerve fiber (IENF) of mice hind paw, which is known to be easily influenced by VIPN. On the foot pad of mice at the 16 days after VCR injection, pathological decrease in number of IENF was shown in both groups, but no difference was observed between the groups (Fig. 2D, E). Taken together, these results show that T. spiralis infection did not alter axonal degeneration induced by VCR.
Fewer infiltrated macrophages were found in the sciatic nerves of T. spiralis infected mice
We hypothesized that the difference in mechanical allodynia between the infected and non-infected groups is due to some differences in inflammation. The axons were not changed significantly in their number and shape despite the VCR treatment by microscopic observation (data not shown), which is consistent with the results of the nerve conduction study. However, we recognized that a certain number of macrophages were infiltrated into the sciatic nerve in response to the VCR treatment. Of note that, fewer infiltrated macrophages were observed in the sciatic nerve of the T. spiralis infected mice than in the non-infected group (Fig. 3A, B). Western blot experiments of sciatic nerves from T. spiralis infected mice also confirmed decreased band intensities of CD68 antigen which reflects reduced macrophage.
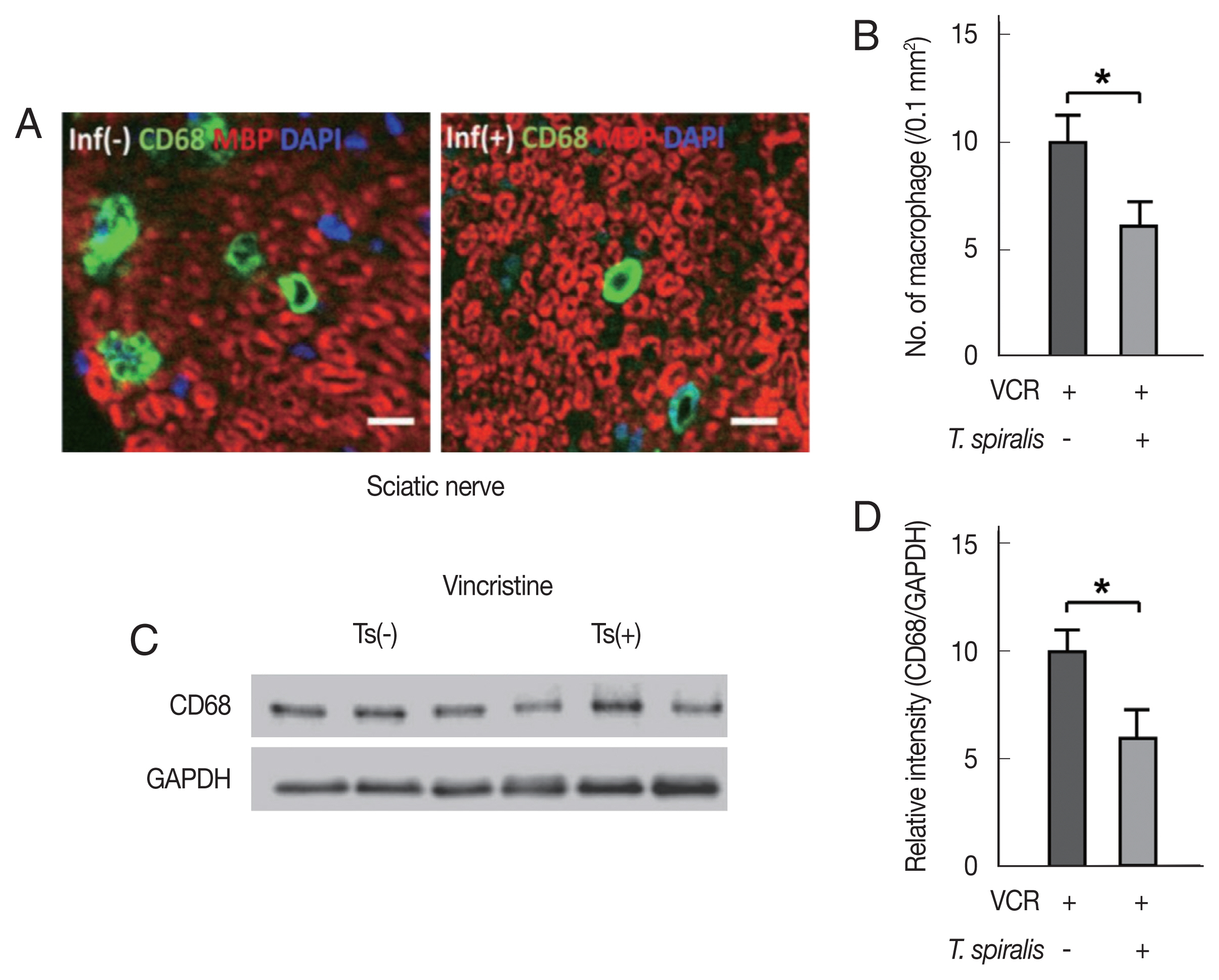
Vincristine (VCR) treatment resulted in decreased the number of infiltrated macrophages in sciatic nerve of Trichinella spiralis infected mice. (A) Representative immunofluorescent images of macrophages (CD68) and myelin (MBP) in the sciatic nerves from uninfected and T. spiralis infected mice. (B) The number of infiltrated macrophages in sciatic nerve sections. Ts; T. spiralis infected group. (C) Western blot analysis showing the expression levels of CD68 in the sciatic nerve after VCR treatment. (D) Mean levels of the band intensity of CD68 normalized to GAPDH.
T. spiralis ESP-treated macrophages secrete reduced amount of IL-1β
Since the T. spiralis probably control the immunity of a distant place through its ESP, we prepared ESP of T. spiralis [17]. We investigated how T. spiralis ESP affected activation of macrophages. The secretion of typical pro-inflammatory cytokines in peritoneal macrophages was also measured by ELISA.
Peritoneal macrophages were obtained from the wild-type mouse and seeded on the 24-well plate, and incubated for 2 h at 37°C, then unattached cells were washed with pre-warmed media. After pre-treatment of ESP (100 μg/ml) for 30 min, LPS (1 μg/ml) was re-treated to stimulate peritoneal macrophages. The supernatant, which was incubated for 24 h, was collected, and ELISA was performed. IL-1β showed a statistically significant decrease (49.7 vs 16.6 pg/ml, P<0.05) in the group that pre-treated with ESP, while IL-6 (3.8 ng/ml vs 3.8 ng/ml) and TNF-α (1.3 ng/ml vs 1.2 ng/ml) showed no statistical differences between groups.
DISCUSSION
In this study, we showed that VCR-induced mechanical allodynia is improved in T. spiralis-infected mice at 10 days after VCR treatment (Fig. 1A). This improvement appears not to be caused by direct inhibition of axonal degeneration, because difference in number of nerve fibers is not observed (Fig. 2). Since decrease of macrophage infiltration in the PNS can improve VCR-induced mechanical allodynia [4], we next investigated number of infiltrated macrophages in sciatic nerves of the T. spiralis-infected mice. We found that the number of infiltrated macrophages in the sciatic nerves of VCR treated mice is reduced in the T. spiralis infected group compared to uninfected group (Fig. 3).
Although the mechanism how VCR increases the number of macrophages in sciatic nerve is not fully understood, VCR treatment is known to trigger Sarm1 activation and Wallerian-like degeneration, similar to those observed by axonal injury [3]. When the Wallerian degeneration occurs, macrophages are recruited in response to chemoattract factors from the damaged nerve [21]. In this study, axon and myelin are not frequently observed in area occupied by the macrophages (Fig. 3A), indicating that macrophages are responsible for phagocytic clearance of injured axon and myelin. The detailed mechanism how T. spiralis infection reduces macrophage infiltration remains to be elucidated, since no significant difference was observed in axonal degeneration in T. spiralis infected mice.
It is known that T. spiralis infection also affect macrophage activities [14,15,22]. Macrophage has ability to secret high level of pro-inflammatory cytokines including IL-1β. The importance of IL-1β in increase of neuron excitability and susceptibility to pain is well established [23,24]. A recent study reported that cystatin derived from T. spiralis suppresses IL-1β and TNF-α secretion of bone marrow-derived macrophage [22]. Similar to the study, we also found that ESP of T. spiralis can inhibit IL-1β secretion of LPS-stimulated mouse peritoneal macrophage (Fig. 4A–C). However, we could not find differences in the secreted IL-6 or TNF-α levels, which may be due to difference in concentration of effector molecules, because the aforementioned study used purified recombinant protein.
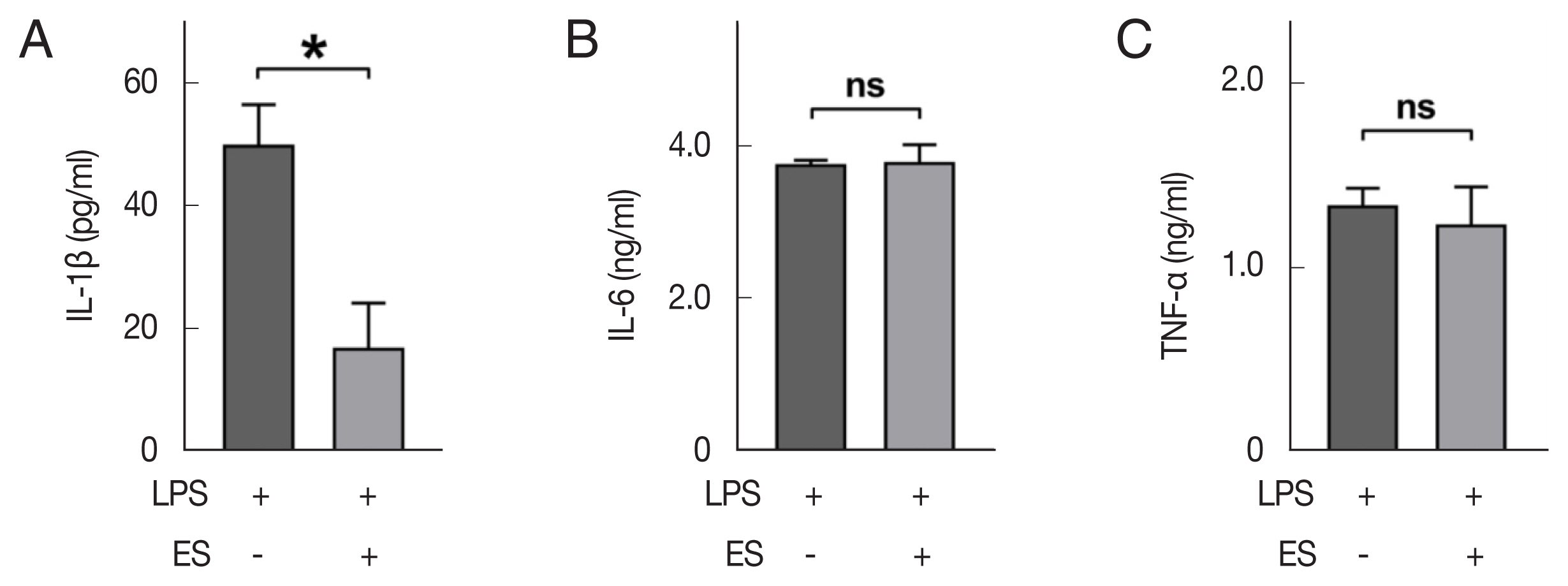
The levels of pro-inflammatory cytokines secreted by LPS-stimulated peritoneal macrophage assessed by ELISA. The peritoneal macrophages were untreated or pre-treated with ES product, followed by LPS (1 μg/ml) for 24 h, after which the culture supernatants were subjected to the ELISA. Comparison of secreted IL-1β (A), IL-6 (B), and TNF (C) between ES pre-treatment and non-treatment groups. Mean±SD (n=3). ES, ES product from Trichinella spiralis. *P<0.05. ns, not-significant.
In addition to the importance of IL-1β in pain, macrophage-derived IL-1β plays a key role in myeloid cell recruitment [25]. It was reported that functional blocking of IL-1β reduces number of recruited macrophages at sciatic nerves of mice during Wallerian degeneration [26]. Taken together, our study suggests that T. spiralis infection may contribute to the alleviation of VCR-induced allodynia by reducing the number of infiltrated macrophages as well as the secretion of inflammatory cytokines.
In the present study, statistical significance of difference in allodynia score was not shown at 15 days after VCR treatment. The reason for this result may be that improvement of inflammation by T. spiralis infection cannot prevent axonal damage itself because VCR-induced axon damage precedes inflammation. In addition, the axonal injury may be greater because the VCR was administered at a higher dose than those applied in humans.
Considering that the use of anti-inflammatory agents in cancer therapy is known to be beneficial [27], T. spiralis can be an appropriate candidate for preventing or treating painful neuropathy accompanying cancer therapy. However, there are some limitations on administrating live T. spiralis to humans. For example, infection of the T. spiralis in humans can provoke several symptoms including diffuse myalgia or fever [28]. In addition, since T. spiralis and ESP of T. spiralis interact simultaneously with many types of cells, there may be therapeutic limitation in that the exact mechanism is unclear. Further studies may need improvement of methods such as purification of effective substances.
ACKNOWLEDGMENTS
This study was supported by grants from the National Research Foundation of Korea (NRF; 2016R1A5A2007009). This research was supported by grant funded by the Ministry of Education (2022R11A1A01072033).
Notes
The authors declare no competing interests.