Usefulness of the recombinant liver stage antigen-3 for an early serodiagnosis of Plasmodium falciparum infection
Article information
Abstract
In order to develop tools for an early serodiagnosis of Plasmodium falciparum infection, we evaluated the usefulness of P. falciparum liver stage antigen-3 (LSA-3) as a serodiagnostic antigen. A portion of LSA-3 gene was cloned, and its recombinant protein (rLSA-3) was expressed in Escherichia coli and purified by column chromatography. The purified rLSA-3 and 120 test blood/serum samples collected from inhabitants in malaria-endemic areas of Mandalay, Myanmar were used for this study. In microscopic examinations of blood samples, P. falciparum positive rate was 39.1% (47/120) in thin smear trials, and 33.3% (40/120) in thick smear trials. Although the positive rate associated with the rLSA-3 (30.8%) was lower than that of the blood stage antigens (70.8%), rLSA-3 based enzyme-linked immunosorbent assay could detect 12 seropositive cases (10.0%), in which blood stage antigens were not detected. These results indicate that the LSA-3 is a useful antigen for an early serodiagnosis of P. falciparum infection.
INTRODUCTION
Malaria, caused by the apicomplexan protozoa Plasmodium sp., is one of the most important infectious diseases worldwide, and the problem appears to be worsening (Breman et al., 2001; Snow et al., 2001). Although the morbidity and mortality rates have been declining gradually, malaria continues to constitute the major cause of mortality and morbidity in many parts of the world. Approximately 300 to 500 million new malaria cases and 2 to 5 million deaths are estimated to occur annually. The emergence and rapid spread of resistant strains to commonly used antimalarial drugs, including chloroquine and antifolates, poses a serious challenge to the malaria control (Wellems and Plowe, 2001; Roper et al., 2003; Baird, 2004; Gregson and Plowe, 2005).
Vivax malaria reemerged in 1993 and continues to be a big health threat to Korea (Chai et al., 1994; Cho et al., 1994; Yeom et al., 2005). Recent increases of overseas travel and economic activities with other countries, especially those having malaria-endemic regions, make another concern with falciparum malaria. In deed, 476 imported malaria cases have been reported from 1994 to 2004 in the Republic of Korea (unpublished data, Korea Center for Disease Control and Prevention). Most of these patients were infected with Plasmodium falciparum and had been infected principally in Asia and Africa where drug resistance strains are prevalent. Therefore, an early diagnosis, with prompt treatment, is highly important. In this respect, development of protocols and materials for an earlier diagnosis of falciparum malaria is urgently necessitated.
The sporozoites of malaria parasites transmitted from the saliva of infected mosquitoes travel rapidly to the liver and invade hepatocytes, where they develop into the exoerythrocytic stage called a tissue schizont. During this stage, the parasites express liver stage specific antigens. In P. falciparum, at least 2 of the relevant antigens, liver stage antigen-1 (LSA-1) and liver stage antigen-3 (LSA-3), have been identified and characterized (Guerin-Marchand et al., 1987; Aidoo et al., 2000; Joshi et al., 2000). These proteins are both surface proteins, and are expressed solely in infected hepatocytes and thought to have a role in liver schizogony and merozoite release. Specific humoral, cellular, and cytokine immune responses to LSA-1 and LSA-3 are well documented, with epitopes identified that correlate with the antibody production, proliferative T-cell responses, or cytokine induction (Aidoo et al., 2000; Joshi et al., 2000; Perlaza et al., 2001). Both preerythrocyte antigens have been considered to be vaccine candidates against P. falciparum due to their antigenic and protection-inducing immunogenic properties (Daubersies et al., 2000; Kurtis et al., 2001; Sauzet et al., 2001; Taylor-Robinson, 2003). In the present study, we evaluated the usefulness of P. falciparum LSA-3 as a serodiagnostic antigen for a more rapid diagnosis of falciparum malaria than using the currently available diagnostic tools.
MATERIALS AND METHODS
Blood samples
Blood samples were collected from 120 inhabitants who visited the malaria clinic with signs and symptoms suggestive of malaria in endemic areas of Mandalay, Myanmar. Thin and thick blood smears were prepared for routine microscopic examinations. Informed consent was obtained from all patients. The study protocol was approved by the Department of Health (Upper Myanmar), the Union of Myanmar.
Cloning and sequence analysis of liver stage antigen-3 (LSA-3) of P. falciparum
To amplify a portion of P. falciparum LSA-3 gene, we designed specific primer sets based on the DNA sequences provided by the Genebank database. LSA-3F contained a BamHI site at its 5' end (5'-GATCCGAAAATGTAGAAGAAAATGACGACGGAAGT-3'), and LSA-3R contained a HindIII site at its 5' end (5'-AAGCTTTGATAAATTTGTTGCAACGTTTTCAGCAAC-3'). Genomic DNA of P. falciparum was extracted from the whole blood of a patient using a QIAamp DNA Blood Kit (Qiagen, Valencia, California, U.S.A.) following the manufacturer's instruction. Polymerase chain reaction (PCR) was performed with AccuPower™ PCR Premix (Bioneer, Daejeon, Korea), 50 ng of purified genomic DNA, and 40 pmoles of the above-described reverse and forward primer sets, in a total volume adjusted to 30 µl with distilled water. The reaction condition was as follows: denaturation at 94℃ for 10 min, 35 cycles of 1 min at 94℃, 1 min at 60℃, and 1.5 min at 72℃, and a final extension for 10 min at 72℃. The PCR product was gel-purified using a Gel extraction kit (Qiagen), ligated into pCR2.1-TOPO™ cloning vector (Invitrogen, Carlsbad, California, U.S.A.) and then transformed into E. coli TOP10. Sequencing reactions were conducted with a BigDye Terminator Cycle Sequencing Ready Reaction Kit in an ABI 377 automatic DNA sequencer (Applied Biosystems, Foster City, California, U.S.A.). Nucleotide and deduced amino acid sequences were analyzed with the SeqEd.V1.0.3 program, and the CLUSTAL program provided in the Megalign software, a multiple-alignment program of the DNASTAR package (DNASTAR, Madison, Wisconsin, U.S.A.).
Expression and purification of recombinant LSA-3 (rLSA-3)
For the expression of P. falciparum LSA-3 in Escherichia coli, the amplified PCR product was digested with BamHI and HindIII and purified with a Gel extraction kit (Qiagen), ligated into pQE30 expression vector (Qiagen), and transformed into E. coli M15 (pREP4) cells (Qiagen). The selected clones were grown and induced with 1 mM isopropyl-1-thio-β-D-galactopyranoside (IPTG). The bacteria were then suspended in native lysis buffer (50 mM NaH2PO4, 300 mM NaCl, 10 mM imidazole, pH 8.0), sonicated on ice and centrifuged at 4℃ for 20 min at 12,000 g. The supernatant was collected and analyzed by SDS-PAGE. The rLSA-3 was purified by nickel-nitrilotriacetic acid (Ni-NTA) chromatography (Qiagen).
Western blot analysis
The purified rLSA-3 was run on 10% SDS-PAGE gel, and the protein was transferred onto a nitrocellulose membrane. After the transfer, the membrane was cut into strips and blocked for nonspecific binding with 3% skim milk for 12 hr at 4℃. The strips were then washed 3 times with phosphate buffered saline (PBS) with 0.15% Tween 20 (PBST) for 10 min each. The strips were allowed to react with the serum samples obtained from either normal or P. falciparum-infected persons (diluted 1:100, v/v) for 4 hr, and then washed 3 times with PBST as described above. Diluted peroxidase-conjugated anti-human IgG (1:1,000, v/v; Sigma, St. Louis, Missouri, U.S.A.) was added to each strip followed by 3 hr incubation at room temperature. For color development, 0.2% diaminobenzidine and 0.002% H2O2/PBS were used.
Enzyme-linked immunosorbent assay (ELISA)
The 96-well polystyrene plates were coated with 1 µg/ml of purified rLSA-3 diluted in PBS (pH 7.4) per well. The plates were incubated overnight at 4℃. After incubation, the plates were washed 3 times with PBST and blocked via 2 hr incubation with 200 µl of 3% BSA in PBST per well for 2 hr at 37℃. After 3 washes with PBST, 100 µl of sera (1:100, v/v) was added to each well, and incubated for an additional 2 hr at 37℃. After washing as described above, the plates were incubated for 2 hr at 37℃, with 100 µl of peroxidase-conjugated anti-human IgG (1:1,000, v/v, Sigma). The plates were then washed 3 times with PBST and 100 µl of substrate solution was added. The substrate solution was prepared immediately prior to use by dissolving 30 mg of o-phenylenediamine (Sigma) per 60 ml of peroxidase solution B (H2O2 in 0.1 M phosphate citric acid buffer, pH 5.0). The plates were then incubated for an additional 30 min at dark, and the reactions were terminated by adding 50 µl of 4 N H2SO4. Optical densities were measured at 450 nm with an ELISA reader.
Immunofluorescent antibody test (IFAT)
IFAT was conducted with smeared P. falciparum-infected red blood cells (blood stage antigens; BSA) on multispot slides. The infected red blood cells were routinely obtained by in vitro cultivation of the parasite in our laboratory. The parasite-infected red blood cells in slides were allowed to react with the test sera (1:32, v/v) for 30 min, then rinsed twice with PBS, and incubated for 30 min with fluorescein isothiocyanate (FITC)-conjugated anti human IgG (1:100, v/v, Sigma). The slides were then observed under a fluorescence microscope.
RESULTS
Cloning of LSA-3 from P. falciparum genomic DNA
The genomic DNA used for PCR amplification of LSA-3 was prepared from a Myanmar patient. Sequencing analysis of the cloned LSA-3 gene was determined to encompass 1,500 bp and encoded for 500 amino acids (Fig. 1). It exhibited a specific amino acid tandem repeats, 27 repeats of VEESVAEN, and 6 repeats of VEEIVAPTVEEI and VAPTVEEIVAPSVVESVAPSVEESVEEN. These repeats are highly conserved in different strains of P. falciparum and the LSA-3 gene from the Myanmar isolate evidenced its highest degree of homology (99.6%) with a Kenya isolate (AJ007010) (data not shown).
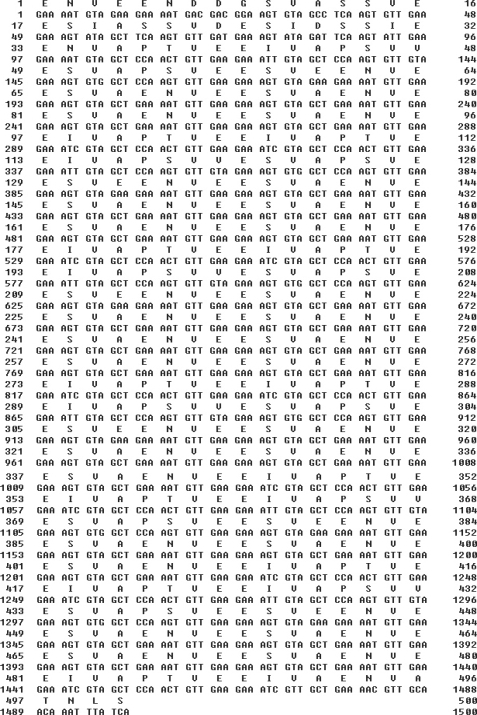
Nucleotide and deduced amino acid sequences of the liver stage antigen-3 (LSA-3) of Plasmodium falciparum Myanmar isolates. Specific amino acid tandem repeats are observed in the sequence.
Expression and purification of recombinant LSA-3 (rLSA-3)
The expression of the rLSA3 was induced by adding IPTG and analyzed on SDS-PAGE. The size of the expressed protein, rLSA3, was approximately 52 kDa, which is consistent with the predicted molecular weight of the protein (Fig. 2A). In the western blot analysis of the purified rLSA-3, 5 serum samples from 14 malaria patients infected with P. falciparum were shown to be reactive against rLSA-3 (Fig. 2B).
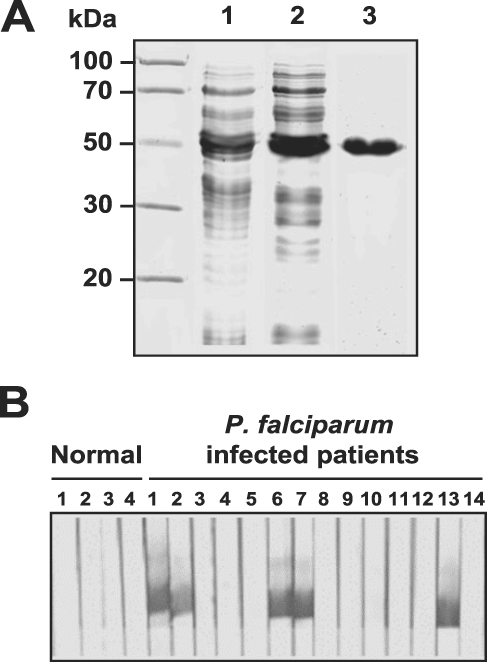
Expression and purification of recombinant LSA-3. (A) SDS-PAGE analysis. Lane 1, uninduced E. coli lysate; lane 2, IPTG induced E. coli lysate; lane 3, Ni-NTA affinity purified rLSA-3. (B) Western blot analysis. Each strip was incubated with sera from P. falciparum-infected patients. Some of the sera (5 of 14 samples) exhibited positive reactions.
Comparison of parasite detection rates
In order to evaluate the utility of rLSA-3 as a serodiagnostic antigen, we conducted several diagnostic tests and the parasite detection rate of each test was compared. In the case of microscopic examinations of the 120 blood samples collected in malaria-endemic areas of Mandalay, Myanmar, the positive rate was 39.2% (47/120) in thin smear trials and 33.3% (40/120) in thick smear trials (Table 1). Cases of mixed infection with P. vivax and P. falciparum were also detected in thin blood smear tests. IFAT employing the whole BSA revealed that 85 (70.8%) among 120 sera tested were determined to have antibodies against BSA. Meanwhile, ELISA employing the rLSA-3, positive rate was lower than that for blood stage antigens, at 30.8% (37/120). However, rLSA-3 based assays could detect 12 seropositive cases (10.0%), in which BSA were not detected (Table 2).

Microscopic examinations of 120 blood samples using thick and thin smears
Comparison of the antibody positive rates of 120 sera against liver stage antigen-3 (LSA-3) and blood stage antigen (BSA)
DISCUSSION
In the present study, we attempted to develop materials through which malaria parasites could be detected prior to the clinical onset of the disease. All human malaria including P. falciparum, remain for several weeks or months in liver cells after their introduction into the human body by mosquitoes, in the form of sporozoites. If the disease could be diagnosed in patients during these pre-onset periods of incubation, chances of early treatment might be greatly improved.
The infected sporozoites of malaria parasites developed and matured in the hepatocytes. During this incubation period, the parasites express liver stage-specific antigens, including LSA-1 and LSA-3. These preerythrocyte antigens evoked specific humoral and cellular immune responses in the hosts and have been recognized as candidates for vaccine (Aidoo et al., 2000; Daubersies et al., 2000; Joshi et al., 2000; Kurtis et al., 2001; Perlaza et al., 2001; Sauzet et al., 2001; Taylor-Robinson, 2003). In this consideration, the liver stage-specific antigen could also be a good diagnostic candidate for early diagnosis of malaria.
In order to determine the feasibility of this, we evaluated the rLSA-3 as a promising candidate marker for screening of patients with liver-resident parasites. As we expected, the antibodies against LSA-3 were proved to be detectable in the patient's sera prior to the production of antibodies specific for BSA. Although positive rate of rLSA-3 based ELISA was lower than that of BSA, rLSA-3 based assay could detect 12 seropositive cases (10.0%), which were not detected by the BSA. Therefore, LSA-3 appears supplemental to the BSA, and may be proved useful as a serodiagnostic adjunct. A larger prospective study with a larger number of samples and different infectious status of samples are needed to generalize this method for a diagnostic use.
ACKNOWLEDGMENTS
This work was supported by the intramural grant of the National Institute of Health, Korea (NIH-347-6111-155/158) and grant from the Korea Health 21 R & D Project, Ministry of Health and Welfare, Republic of Korea (03-PJ1-PG1-CH01-0001).