Heterophyid metacercarial infections in brackish water fishes from Jinju-man (Bay), Kyongsangnam-do, Korea
Article information
Abstract
Heterophyid metacercarial infections in brackish water fishes, i.e., perch, shad, mullet, redlip mullet, and goby, of Jinju-man (Bay), Kyongsangnam-do, Korea, were investigated using a digestion technique. Among 45 perch (Lateolabrax japonicus), the metacercariae of Heterophyopsis continua were found in 55.6% (18.5 metacercariae per fish), Stictodora spp. in 28.9% (3.6), and Metagonimus takahashii in 6.7% (17.0). The metacercariae of H. continua were detected in 23 (65.7%) of 35 shad (Konosirus punctatus). Among 15 mullet (Mugil cephalus), the metacercariae of Pygidiopsis summa were found in 100% (105.9 metacercariae per fish), Heterophyes nocens in 40.0% (8.5), H. continua in 13.3%, and Stictodora spp. in 6.7%. Among 12 redlip mullet (Chelon haematocheilus), the metacercariae of P. summa were detected in 91.7% (1,299 metacercariae per fish), H. nocens in 16.7%, and Stictodora spp. in 16.7%. Among 35 gobies (Acanthogobius flavimanus), the metacercariae of Stictodora spp. were found in 82.9% (44.5 metacercariae per fish), and H. continua in 25.7% (2.8). The adult flukes recovered from a cat experimentally infected with metacercariae from perches were compatible with M. takahashii. The results confirm that the estuarine fish in Jinju-man (Bay) are heavily infected with heterophyid metacercariae. Furthermore, it was found for the first time that the perch acts as a second intermediate host for M. takahashii.
INTRODUCTION
About 30 species of 9 genera, i.e., Metagonimus, Heterophyes, Heterophyopsis, Stellantchasmus, Centrocestus, Pygidiopsis, Procerovum, Haplorchis, and Stictodora, are known to be human-infecting heterophyid flukes (Chai and Lee, 2002). During the 1970's it was believed that Metagonimus yokogawai was the only human pathogenic fluke in Korea. However, since then, additional human pathogens, such as Heterophyes nocens, Heterophyopsis continua, Stellantchasmus falcatus, Pygidiopsis summa, Centrocestus armatus, Metagonimus takahashii, Metagonimus miyatai, Stictodora fuscata and Stictodora lari have been added to the list of pathogenic heterophyid flukes (Chai and Lee, 2002). Nevertheless, considerably earlier, trematodes of genus Heterophyes were suggested to have a national distribution and possibly infect man, after eggs were discovered in the human feces (Stryker, 1914; Imanishi, 1914), metacercariae in mullet (Asada, 1934), and worms from animal intestines (Kobayashi, 1925; Furuyama, 1930).
Heterophyid flukes can cause abdominal pain and diarrhea in infected humans, and their eggs, after being transferred by the blood stream, may produce eosinophilic granuloma in the heart, brain, and spinal cord (Africa et al., 1940; Kang et al., 1983; Cho et al., 1985). In terms of sources of infection, several kinds of brackish water fish, such as, perch (Lateolabrax japonicus), shad (Konosirus punctatus), mullet (Mugil cephalus), and goby (Acanthogobius flavimanus), have been identified in Korea (Chun 1960, 1963; Seo et al., 1980, 1981, 1984; Chai and Sohn, 1988; Chai et al., 1989; Sohn et al., 1994; Sohn and Moon, 2001). These fish are sometimes heavily infected with the metacercariae of heterophyid flukes, and are popularly eaten raw by residents in coastal areas. However, no simultaneous investigation has been conducted on the infection status of heterophyid metacercariae in a range of estuarine fish species, in a particular area. Therefore, the purpose of this study is to systematically investigate the infection status of heterophyid fluke metacercariae in estuarine fish species caught in Jinju-man (Bay), which is located in the middle of southern coast of Korea.
MATERIALS AND METHODS
Between early September and late October of 2004, 45 perch (L. japonicus), 35 shad (K. punctatus), 15 mullet (M. cephalus), 12 redlip mullet (Chelon haematocheilus), and 35 gobies (A. flavimanus) were collected from Jinju-man in Kyongsangnam-do. All collected fishes were transferred to our laboratory, measured for length and weight, and examined by artificial digestion method (Table 1). Trematode metacercariae were segregated, collected under a stereomicroscope, and categorized according to measurements and morphological characteristics. Intensities of infection and infection rates were then calculated. Metagonimus sp. metacercariae were used to experimentally infect a cat to obtain adult worms. Twenty-one days after infection, adult worms were recovered from the small intestine, fixed with 10% neutral buffered formalin, and stained with Semichon's acetocarmine in order to study their morphological characteristics.

Estuarine fish species examined in this study
RESULTS
Infection status of heterophyid metacercariae
Metacercariae of H. continua, Stictodora spp. and M. takahashii were detected in the perch (Table 2). Total 152 metacercariae of H. continua (1-40/fish) were detected in 23 (65.7%) of 35 shad. The metacercariae of P. summa, H. nocens, H. continua and Stictodora sp. were detected in mullet (Table 3). In redlip mullet (Fig. 1), we detected the metacercariae of P. summa, H. nocens and Stictodora sp. (Table 4), and in gobies, we detected the metacercariae of Stictodora spp. and H. continua. In total, 1,291 metacercariae of Stictodora spp. (1-194/fish) were collected from 29 gobies (82.9%). H. continua metacercariae were from 9 gobies (27.7%), with an average infection density per infected fish of 2.8 metacercariae (1-8/fish).
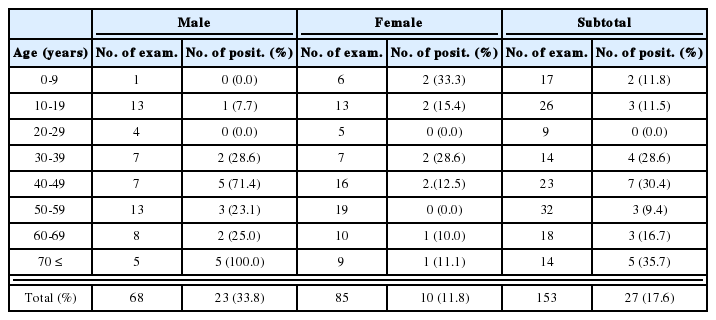
Infection status of heterophyid metacercariae in the perch, Lateolabrax japonicus, from Jinju-man
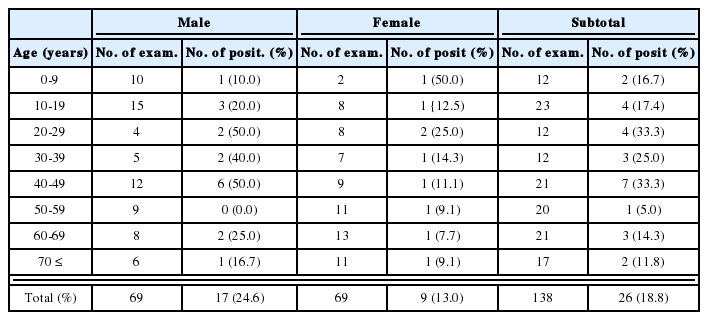
Infection status of heterophyid metacercariae in the mullet, Mugil cephalus, from Jinju-man

Mullet, Mugil cephalus (upper) and redlip mullet, Chelon haematocheilus (lower). The redlip mullet has been shown to act as a second intermediate host for heterophyid flukes, i.e., H. nocens, P. summa and Stictodora sp., in the present study.
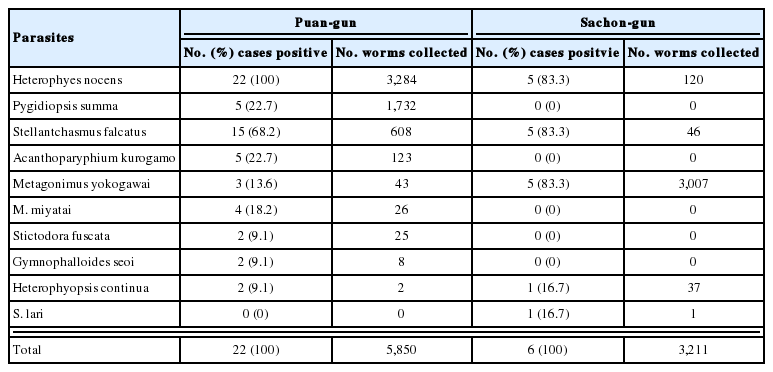
Infection status of heterophyid metacercariae in the redlip mullet, Chelon haematocheilus, from Jinju-man
Morphology of metacercariae and adults of Metagonimus takahashii (Fig. 2)

The new second intermediate host, Lateolabrax japonicus (A) (bar = 30 mm), a metacercaria (B) (bar = 0.05 mm), and an adult (C) (bar = 0.25 mm) of Metagonimus takahashii. Metacercariae were characterized by a disc shape, ventral sucker displaced from the median, yellow-brownish pigments scattered in the body, and excretory bladder (EB) flat and V-shaped. Adults were recovered from a cat experimentally infected with the metacercariae, and characterized by a small-sized, rightwardly displaced ventral sucker (larger than the oral sucker), the uterine loop passed between 2 testes (T) and vitellaria (arrows) distributed over the posterior end of the right testis.
The disc shaped metacercariae of M. takahashii detected in the perch were 150-175 (mean 162) x 150-163 (mean 156) µm in size. Their ventral suckers, deviated to one side and were larger than the oral sucker. Brownish pigments were scattered over whole bodies and a black excretory bladder was located near the posterior end. Adults recovered from an experimental cat were 580-820 (mean 688) x 250-350 (mean 288) µm in size, and had a sharp anterior and a round posterior end. A round oral sucker (mean 39 x 53 µm) was located at the anterior end, followed by a muscular pharynx (mean 31 x 24 µm), and a relatively long esophagus (mean 93 µm) and the intestine. The right deviated ventral sucker (mean 77 x 49 µm) was located at the anterior 2/5 of worm. The oval-shaped ovary (mean 61 x 59 µm) was located in posterior 1/3-1/2 median. The seminal vesicle (mean 65 x 147 µm) was long saccular, often divided into two parts by a constriction (mean 59 x 35 µm), and was transversely located in the left space between the ventral sucker and ovary. The seminal receptacle (mean 116 x 92 µm) was irregular in shape, and situated near the right side of the ovary. Two testes were arranged obliquely in tandem and apart from each other. The right testis (mean 159 x 88 µm) was located more posteriorly than the left testis (mean 135 x 92 µm). The uterus occupied all available space from the acetabular level to near posterior end of the body, and the uterine loop passed between the two testes. Vitellaria extended along the right lateral side of the body and reached over the posterior end of the right testis. Uterine eggs were ovoid, light yellowish brown in color, and 28.8-32.5 µm (mean 31.0 µm) x 17.5-20.0 µm (mean 18.6 µm) in size.
DISCUSSION
Investigations of the infection rates of metacercariae in second intermediate hosts, in combination with studies on adult worm infection rates in humans and larval infections in first intermediate hosts, can usefully describe the epidemiology of trematodes in a particular area. However, in the case of heterophyid flukes, stool examinations are not suitable in order to understand infection rates in humans and reservoir hosts, since the amount of egg production is too small, and the first intermediate hosts are not well studied. Therefore, in order to provide epidemiologic information on heterophyid trematodes, we investigated the infection status of metacercariae in estuarine fishes, namely, perch, shad, mullet, redlip mullet, and gobies that were caught in Jinju-man (Bay), Kyongsangnam-do, which is located in the middle of southern coast of Korea.
Several workers have reported on heterophyid fluke metacercariae collected from estuarine fish species in Korea (Chai and Lee, 2002). Chun (1960) detected H. continua metacercariae in perch, shad, and gobies from Kyongsangnam-do. Chun (1963) first isolated P. summa metacercariae from mullet. Seo et al. (1980) newly described the metacercariae of H. nocens from mullet. Chai and Sohn (1988) identified S. falcatus metacercariae encysted in mullet. Chai et al. (1989) detected S. lari metacercariae in gobies, and Sohn et al. (1994) found S. fuscata metacercariae in gobies. Sohn et al. (2003) also identified Acanthotrema felis metacercariae collected from gobies. However, in the present study, the metacercariae of S. falcatus and A. felis were not detected though M. takahashii metacercariae are reported for the first time in perch. Previously, the common carp, Cyprinus carpio, the crussian carp, Carassius auratus, and the dace, Tribolodon taczanowskii, have been reported to be intermediate hosts of M. takahashii (Chai and Lee, 2002). Thus, the present study shows for the first time that the perch, L. japonicus, is a second intermediate host for M. takahashii in Korea.
Many species of fishes, i.e., M. cephalus, L. japonicus, A. flavimanus, K. punctatus, Plecoglossus altivelis, Conger myriaster, Harengula zunasi, Dorosoma thrissa, Coilia sp., Cyprinus carpio, Mugil affinis, Gobius nebulosus, Boleophthalmus pectinirostris and Scartelaos sp. have been reported to be second intermediate hosts for H. continua in Japan, Korea, and China (Kanemitsu et al., 1953; Chun 1960; Komiya, 1965; Kobayashi, 1968; Seo et al., 1984; Cho and Kim, 1985; Sohn et al., 1994; Kim et al., 1996; Sohn and Moon, 2001; Sohn et al., 2005). Among these, M. cephalus, L. japonicus, A. flavimanus, K. punctatus, P. altivelis, C. myriaster, B. pectinirostris and Scartelaos sp. were recorded in Korea. In the present study, H. continua metacercariae were also detected in perch, shad, mullet, and gobies.
Endemic foci of H. nocens infection are scattered in southwestern coastal areas and on offshore islands, especially in Shinan-gun and Muan-gun, Jeollanam-do, Korea (Chai and Lee, 2002). Moreover, some species of estuarine fishs, i.e., M. cephalus, A. flavimanus, Tridentiger obscurus, Liza haematocheila, Therapon oxyrhynchus, Glossogobius giuris brunneus, B. pectinirostris and Scartelaos sp. have been reported to be infected with H. nocens metacercariae in Japan and Korea (Komiya, 1965; Seo et al. 1980, 1981; Sohn et al., 2005). In the present study, H. nocens metacercariae were detected in mullet and redlip mullet. The fact that the redlip mullet, C. haematocheilus, serves as a second intermediate host for H. nocens, was also confirmed by the present study, and thus, the redlip mullet is presumed to act as the source of human infections in endemic areas. Moreover, L. haematocheila previously reported as a second intermediate host for H. nocens in Japan is probably a synonym of the redlip mullet, C. haematocheilus.
Since the first discovery of P. summa metacercariae by Chun (1963) in Korea, some workers have reported on their infection status in fish hosts, i.e., in mullet and gobies (Seo et al., 1981; Sohn et al., 1989 & 1994; Sohn and Moon, 2001). Seo et al. (1981) detected 15,230 metacercariae in 17 mullet from 4 southwestern coastal areas, and found 9 metacercariae in only one goby from Mokpo-shi, Jeollanam-do. Sohn et al. (1989) collected a total of 7,725 metacercariae from 5 mullet in Samcheonpo-shi, Kyongsangnam-do. Sohn et al. (1994) also detected 31 metacercariae in 10 mullet from Haenam-gun, Jeollanam-do, and Sohn and Moon (2001) found 3,822 metacercariae in 23 (32.9%) of 70 mullet examined. In the present study, we collected an average of 1,299 and 106 of P. summa metacercariae from 15 (100%) mullet and 11 (91.7%) redlip mullet, respectively, however, they were not found in gobies. Thus, the present study reveals the redlip mullet as a second intermediate host and a likely source of human infections.
The morphology of the ventrogenital sac of genus Stictodora flukes plays an important role in species categorization, especially in terms of the number and arrangement of chitinous spines. In the present study, the Stictodora spp. metacercariae detected were S. lari and S. fuscata, which have been previously reported (Chai et al., 1989; Sohn et al., 1994a, b). Sohn et al. (2003) also found A. felis metacercariae in gobies from Haenam-gun. Nevertheless, the present study simultaneously detected Stictodora spp. metacercariae in gobies, perch, mullet and redlip mullet. Moreover, our study demonstrates that the perch and redlip mullet act as second intermediate hosts for Stictodora spp.
The finding that the redlip mullet acts as a second intermediate host for some heterophyid flukes, H. nocens, P. summa and Stictodora sp., is novel. We believe that previous studies have probably not differentiated metacercariae in mullet and redlip mullet due to their morphological similarities (Fig. 1).
Three species of Metagonimus flukes, i.e., M. yokogawai, M. takahashii and M. miyatai are known to be distributed in Korea. They share a larger ventral sucker, deviated to the right, which is larger than the oral sucker, but can be differentiated by uterus location. The uterine loop passes through the anterior border of both testes in M. yokogawai, whereas it courses between both testes in M. takahashii and M. miyatai. The main distinguishing characteristics of M. takahashii and M. miyatai are egg sizes and the distribution of vitellaria. The eggs of M. takahashii are slightly larger than those of M. miyatai, and the vitellaria of M. takahashii are distributed over the posterior end of right testis, whereas in M. miyatai they are located up to 2/3 level of right testis (Saito et al., 1997). The adult worms recovered from a cat experimentally infected with metacercariae obtained from the perch in this study, were morphologically compatible with M. takahashii.
Previous studies and the present study suggest that those who consume raw estuarine fishs, such as perch, shad, mullet, redlip mullet or goby are at a high risk of infection with heterophyid flukes. Moreover, since a proportion of the Korean population eat raw fishs, our findings suggest that the human infection rate may be high. We recommend systematic epidemiological investigations and continuous monitoring, to manage heterophyid infections among populations at a high risk.
Notes
This work was supported by an Anti-Communicable Diseases Control Program of the National Institute of Health (NIH 348-6111-215), Ministry of Health and Welfare, Republic of Korea.