Susceptibility of several strains of mice to Echinostoma hortense infection
Article information
Abstract
Susceptibilities of 5 different mice strains, including C3H/HeN, BALB/c, C57BL6, FvB and ICR, to Echinostoma hortense infection, was evaluated. The worm expulsion rate, worm size and egg production were observed from 1 to 8 weeks after infection with 30 metacercariae. C3H/HeN and ICR mice showed the highest worm maturation rates. The worm recovery rate and the number of eggs per gram (EPG) of feces was also higher in C3H/HeN and ICR mice than in BALB/c, C57BL6, and FvB mice. It is suggested that E. hortense is highly infectious to ICR and C3H/HeN mice, but not to the other strains of mice. Based on the results obtained, we believe that the susceptibility of different mouse strains to E. hortense infection is dependent on the genetic and immunologic background of mice.
INTRODUCTION
Echinostoma spp. are common intestinal parasite in southeast Asia where they have a high prevalence (from 1 to 30%). The presence of this species in Korea was first documented in 1938 (Park, 1938). The first human infection was reported in 1983 by identification of an adult fluke (Seo et al., 1983) and the human cases have been increased gradually. The major symptoms due to Echinostoma infections are abdominal pain, diarrhea, and easy fatigueability. Heavy infections are characterized by severe inflammation and ulceration of the intestinal wall and may cause diarrhea with blood and abdominal pain (Chai and Lee, 2002).
The resistance of the host against intestinal parasites is known to reduce the size and fecundity of the female parasite and the suppress growth during in adult stage (Chai et al., 1984; Ito and Kamiyama, 1987). The variation in host resistance observed is explained by the genetic characteristic of each host (Wakelin et al., 1993). It is presumed that the genetic characteristic of each mouse dictate immune response to parasitic infections.
There are many reports regarding the genetic influence of host in their susceptibility to parasite infections. The difference in susceptibility to Eimeria vermiformis infection among different strains of mice was related to genetic difference in cellular and cytokine response (Wakelin et al., 1993). Strains of mice with different genetic backgrounds exhibited varying susceptibility to Metagonimus yokogawai infection (Chai et al., 1984). But little information has been available concerning immunity against E. hortense.
In this study, we examined the different worm development in different mouse strains of C3H/HeN(H-2k), BALB/c(H-2d), C57BL6(H-2b), FvB(H-2q) and ICR infected with E. hortense experimentally through the detection of worm recovery rate, length and width of several organ and egg lying capacity of the worms.
MATERIALS AND METHODS
Experimental animals
Development and worm recovery of E. hortense in mice: Six-week-old BALB/c, C3H/HeN, C57BL6, FvB and ICR mice were purchased from Daehan Biolink Co. Ltd. (Chungchungbuk-do, Korea), and each strain 40 mice were divided into 8 groups, which were infected with E. hortense and maintained for 8 weeks after infection.
Egg laying capacity of E. hortense in mice: Six-weekold C3H/HeN and ICR mice were divided into each 3 mice, which were infected with E. hortense and maintained for 8 weeks after infection.
Life cycle of E. hortense
We obtained metacercariae of E. hortense from loaches, Misgurnus anguillicaudatus, and used these to infect Sprague-Dawley rats. E. hortense for experimental work was collected from the intestines of these rats 4 weeks later. The E. hortense adults worms collected were washed out using 37℃ PBS (phosphate buffered saline), and spawned for 12 hrs or more. We cultured the eggs in saline at room temperature, and hatched miracidia were infected into the snail (Radix auricularia coreana), which were kept in a glass tank. Cercariae obtained from the snails were infected into M. anguillicaudatus, which were maintained in a glass tank until the cercariae became metacercariae.
Infection of mice and adult worm recovery
We fed 30 E. hortense metacercariae separated from M. anguillicaudatus to each mouse and sacrificed at an interval of 1 week. The intestine was excised, cut open and got the infection rate after confirm the adult worms.
Monitoring of the egg laying capacity
We collected all feces passed over 24 hr periods weekly to determine the egg laying capacity of E. hortense throughout the infection period. EPG (No. of eggs per a gram of feces) was calculated using the formalin-ether concentration and Stoll's egg count methods.
Growth of E. hortense in the host
Adult worms from mouse intestines were examined under the microscope weekly after fixing with 10% formalin, and staining with Semichon's acetocarmine. Worm sizes were obtained using a micrometer and averaged.
RESULTS
Egg laying capacity
EPG were confirmed in C3H/HeN and ICR mice, but not in C57BL6, BALB/c and FvB because eggs were not expelled in the feces. In the case of C3H/HeN, as a result of EPG by Stoll's egg count method, the eggs began to be expelled into the feces 2 weeks after infection. EPG peaked on the 3rd week and then decreased gradually, no more were observed eggs in the feces 6 weeks infection. In the case of ICR mice, eggs first appeared in the feces 2 weeks after infection. The EPG of ICR mice peaked on the 3rd week, and then decreased gradually. No eggs were observed in the feces on the 6th week post-infection (Table 1).

Egg laying capacity of Echinostoma hortense in 6 mice of 2 strains
Growth of E. hortense
Measurements of worms recovered every week after infection, showed that worms in C3H/HeN mice were larger than those from ICR 1 week postinfection, while the worms from ICR were larger than those from C3H/HeN 3 weeks postinfection. In the case of BALB/c mice, the worms were 1.56 ± 2.1 × 0.28 ± 0.02 mm after 1 week and 4.23 ± 2.4 × 0.6 ± 0.02 mm after 3 weeks. Worms from C57BL6 mice were 1.54 ± 0.03 × 0.3 ± 0.02 mm after 1 week and 4.26 ± 15.4 × 0.6 ± 0.02 mm after 3 weeks, and those from FvB mice 1.52 ± 4.28 × 0.3 ± 0.01 mm after 1 week and 4.27 ± 21.2 × 0.56 ± 0.04 mm after 3 weeks. The growth rates in C57BL6, BALB/c and FvB mice were lower than in ICR and C3H/HeN mice (Table 2).
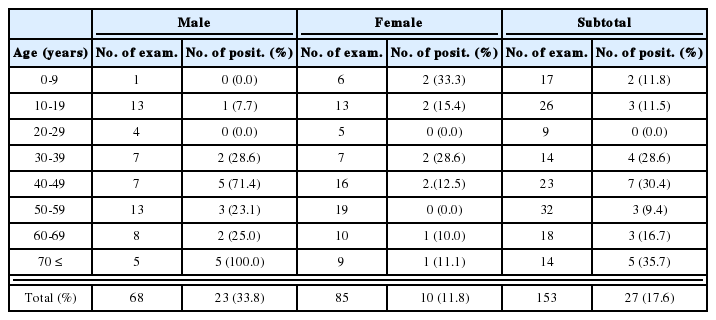
Development of Echinostoma hortense in 200 mice of 5 strains
Recovery rate of worm
The recovery of worms obtained by sacrifice after periods of infection in C3H/HeN mice was continued for 5 weeks after infection, and thus decreased suddenly from the 3rd week. The recovery of worms from ICR mice was continued for 4 weeks, and these also decreased suddenly after 3 weeks of infection. BALB/c, C57BL6 and FvB mice showed worm recovery for 3 weeks after infection, and no worms were found from 4th weeks.
As a result, C3H/HeN and ICR mice were found to give the highest worm recovery rates (Table 3).
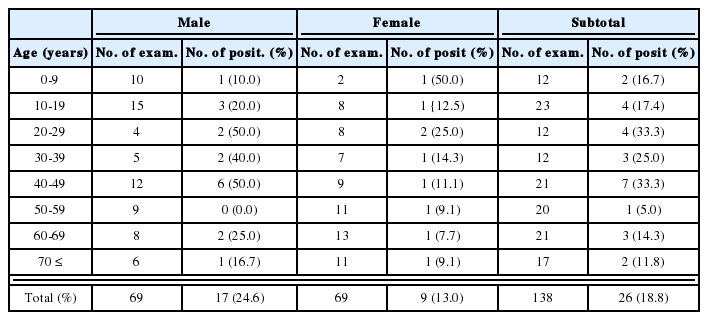
Worm recovery rates (%) after infection with 30 metacercariae in 200 mice of 5 different strains
DISCUSSION
After infecting 5 strains of mice (C3H/HeN, BALB/c, C57BL6, FvB and ICR) with E. hortense, eggs began to be expelled into the feces from two weeks postinfection, and the egg laying capacity of the parasite peaked during the 3rd week after infection. The EPGs of ICR and C3H/HeN mice were 950 and 500 three weeks after infection, respectively, but in the cases of BALB/c, C57BL6 and FvB, no eggs were observed in feces after the 3rd week. Based on the measurements taken, the oral sucker, testis and ovary of parasites in ICR and C3H/HeN mice were larger and developed faster than in the other 3 strains.
According to Lee et al. (1997), who examined the susceptibility of various mouse strains to infection by Gymnophalloides, an intestinal trematode, ddY mice had the largest worms 7 days after infection, next was C3H/HeN, then ICR and BALB/c in order, and KK mice were found to have the smallest worms. By observing the number of eggs in the uterus 7 days after infection, C3H/HeN had the most, then ddY, ICR, KK and BALB/c in order. That is, the susceptibility to G. seoi ran in the order C3H/HeN to ddY to ICR. This result is similar to our result, namely that C3H/HeN and ICR mice are more susceptible than the other strains.
Ahn et al. (1986) reported the EPD of E. hortense in the case of rat infection, when rats were infected with 275 metacercariae, and found that one worm expelled 400-500 eggs per day (400-500 EPD/worm) after 11 days of infection and 1000-1,500 eggs per day after 4 weeks of infection. The egg laying capacity then decreased from 6th week after infection. This result also shows a trend to one found in the present experiment, namely that the egg laying capacity of ICR and C3H/HeN showed an upward tendency until 3 weeks postinfection, but then suddenly decreased and eggs disappeared from six week postinfection.
Kook et al. (1998) reported upon an examination of the egg laying capacity and worm recovery rate of Neodiplostomum seoulense using BALB/cA, C3H/HeJ, F1, mice hybridized BALB/cA with C3H/HeN as a host. All 3 species began to expel eggs 7 days postinfection and this continued until the 21st day, moreover, the worm recovery rate was high in BALB/cA, F1 and C3H/HeJ. Also, the worm survival rate was C3H/HeJ 0%, F1 mouse 26.7% and BALB/cA 86.7%, thus BALB/cA mice were most susceptible to N. seoulense.
We found that the worm recovery rate was 65.7% after one week and 53.3% after two in C3H/HeN, and 66.6 ± 8.25% after one week and 64.7 ± 9.3% after two for ICR mice. So, C3H/HeN and ICR produced higher worm recovery rates than BALB/c, C57BL6 or FvB.
When G. seoi was infected to the various mouse strains, the worm recovery rate from the ICR mouse was highest, next was BALB/c, then KK, C3H/HeN, and ddY in order, C57BL/6 mice showed the lowest recovery rates (Lee et al., 1997). In our study, the ICR mice showed a similar result, whereas C3H/HeN and BALB/c were different susceptibility.
On the worm recovery rates of N. seoulense from various strains of mice, Chai et al. (2000) reported that C57BL/6 produced the highest worm recovery rate followed by C3H/He then C57BL6 among BALB/c (H-2d), C3H/He (H-2k) and C57BL6 (H-2b).
In terms of the time taken to worm expulsion, C3H/He was the earliest and BALB/c the latest, and in terms of the size of worms recovered 7 days after infection, C3H/He produced the smallest followed by BALB/c and C57BL/6 in order, which differed from the results of the present worm size study.
On infecting KK and C3H with Metagonimus yokogawai, the worm recovery rates were KK 18.9% and C3H 1.2% 7 days postinfection, so the result was revealed different from the mouse strain (Chai, 1979; Chai et al., 1984).
It was reported that inflammatory reactions and egg laying capacity are dependent upon mouse genetic differences in Trichinella spiralis infection (Wakelin and Donachie, 1981). It is known that this difference in immunity is the result of differences in the T celldependent immune reaction triggered by the infection (Krco et al., 1983; Else and Grencis, 1991).
Thus, the immunity of hosts infected by parasites depend upon the kinds of parasites and hosts. In the cases of Eimeria vermiformis and Metagonimus yokogawai, the hosts showed different immune responses and it has been reported that genetic differences influence cellular immunity and cytokine expression (Chai et al., 1984; Wakelin et al., 1993). The immune difference in parasite infections is influenced by the differences of immune reaction by parasites and H-2 gene expression (Krco et al., 1983; Else and Grencis, 1991; Wakelin et al., 1993).
In a susceptibility study, some differences in host tolerance are observed for the different strains infected with E. hortense, and it indicates that the different strains of mouse have the different genetic factors.