In vitro culture of Cryptosporidium muris in a human stomach adenocarcinoma cell line
Article information
Abstract
We investigated the optimal culture conditions for Cryptosporidium muris in a human stomach adenocarcinoma (AGS) cell line by determining the effects of medium pH and of selected supplements on the development of C. muris. The optimum pH of the culture medium required for the development of C. muris was determined to be 6.6. The number of parasites significantly increased during cultivation for 72 hr (p < 0.05) at this level. On the other hand, numbers decreased linearly after 24 hr of incubation at pH 7.5. When cultured in different concentrations of serum, C. muris in media containing 5% FBS induced 4-7 times more parasites than in 1% or 10% serum. Of the six medium supplements examined, only 1 mM pyruvate enhanced the number of C. muris in vitro. Transmission electron microscopic observation showed the developmental stages of C. muris in the cytoplasm of the cells, not in an extracytoplasmic location. The growth of C. muris in AGS cells provides a means of investigating its biological characteristics and of testing its response to therapeutic agents. However, a more optimized culture system is needed for the recovery of oocysts on a large scale in vitro.
INTRODUCTION
Cryptosporidium is an intracellular coccidian parasite of humans and many vertebrates worldwide, which is transmitted by oocysts, normally in a fecal to oral manner. In humans, the protozoa infect intestinal epithelial cells, resulting in an acute self-limiting diarrhea in immunocompetent individuals. However, the infection produces more severe and potentially fatal symptoms in immunocompromised individuals (Current and Garcia, 1991).
In the genus Cryptosporidium, at least six species are currently recognized, and C. parvum and C. muris are regarded as mammalian species (Tzipori and Ward, 2002). It has been suggested that C. muris could infect humans, especially immunocompromised patients, because they show a lack of host specificity (Iseki et al., 1989). The oocysts of C. muris were found in the stools of two healthy girls in Indonesia, suggesting the possibility of C. muris infecting humans (Katsumata et al., 2000). Recently, a case of C. muris infection was reported in a HIV-infected adult with diarrhea in Kenya. This was confirmed by both microscopic examination and sequence analysis of the appropriate 840-bp region of the 18S rRNA gene (Gatei et al., 2002).
In vitro culture system of C. parvum has been investigated using numerous cell lines that support the growth of both the sexual and asexual stages of this intracellular parasite (Gut et al., 1991; Yu et al., 2000; Hijjawi et al., 2001). Such in vitro methods are considered to be less expensive and more convenient for the study of host-parasite interactions and for the evaluations of oocyst viabilities and drug sensitivities of the different developmental stages of Cryptosporidium than in vivo models (Gut et al., 1991). Moreover, in vitro models enable the amplification of parasitic material for further immunological, biochemical and molecular studies (Hijjawi et al., 2001). C. muris has been cultured in vitro using a human stomach adenocarcinoma (AGS) cell line (Yu and Choi, 2000), but optimal conditions for an in vitro culture of C. muris needs to be evaluated further in detail. Therefore, we identified optimal culture conditions for C. muris in AGS cells particularly with respect to pH, and examined the effects of various supplements on the development of C. muris in vitro.
MATERIALS AND METHODS
Purification of C. muris oocysts
The oocysts of C. muris were isolated from laboratory mice, which had been maintained in a Parasitology laboratory at Konkuk University, Korea. Five-week old C57BL/6J mice were immunosuppressed with an intramuscular injection of Depomedrol® (0.1 mg/kg, Korea Upjohn Ltd., Korea) one week before C. muris infection. The feces containing oocysts were collected in 2.5% potassium dichromate and kept at 4℃.
The oocysts were isolated using the method described by Petry et al. (1995) with modifications. Briefly, the feces were passed through a metal sieve with a pore size of 500 µm, and the liquid obtained was centrifuged at 1,500 g for 10 min. The pellet was resuspended in saturated NaCl solution, and the uppermost layer containing oocysts was obtained after centrifugation. The oocysts were then resuspended in layered Percoll solution containing two different specific gravities, 1.04 and 1.08, and collected at the interface. Oocysts were kept in 100 U/ml penicillin G, 100 µg/ml streptomycin, and 0.2 mg/ml amphotericin B at 4℃ until required.
Culture conditions and medium supplements
The human stomach adenocarcinoma (AGS) cell line was obtained from the Korean Cell Line Bank, Seoul, Korea. The cells were cultured in RPMI 1640, pH 7.4, with L-glutamine (2 mM) containing N-2-hydroxyethylpiperazine-N'-2-ethanesulfonic acid (HEPES, 20 mM), sodium bicarbonate (24 mM), penicillin G (100 U/ml), and streptomycin (100 µg/ml). For routine cell passage, 10% fetal bovine serum (FBS, Gibco BRL, Gaithersburg, MD, USA) was used. For C. muris infection, the host cells were plated onto a 22 mm2 cover glass, placed in 6-well plates and incubated at 37℃ in a 5% CO2 air humidified incubator.
To investigate if host cell adherence to the surface of the cover glass could enhance the development of C. muris, cover glasses were coated with the cell culture substrates laminin and poly-L-lysine, before infecting with C. muris. The effect of medium pH on in vitro growth of C. muris was also investigated by inoculating excysted oocysts in fresh media, which had been adjusted to the desired pHs using 30 mM each of Tris, MOPS (3-[N-morpholino]propanesulfonic acid) and MES (2-[N-morpholino]ethanesulfonic acid) buffers when the cells reached 70% confluency.
Seven different medium supplements were evaluated for their effects on the in vitro development of C. muris. The supplements examined were; 1, 5 and 10% of fetal bovine serum (FBS); 0.1, 0.2 and 0.4 mg/ml of sodium choleate; 1, 2.5, 4 and 5 mM of pyruvic acid; 8.75, 17.5, and 35 µg/ml of ascorbic acid; 0.25, 0.5, 2, 3, and 4 µg/ml of folic acid; 0.5, 1, 2 and 4 µg/ml of calcium pantothenate; and 1, 4, 8, and 12 µg/ml of 4-(para)-aminobenzoic acid (PABA). All reagents except the FBS, were purchased from Sigma.
Parasite inoculation and the evaluation of infection
Oocysts of C. muris were washed once with RPMI 1640, after excystation, by incubation in 1% trypsin in PBS at 37℃ for 1 hr. They were then inoculated at 1 × 106/well after the host cells had reached 70% confluency. Cover glasses were removed from the wells after 24 hr of incubation, washed three times in PBS (pH 7.4), and fixed in 10% formalin for 30 min. After staining with hematoxylin and eosin, the total number of parasites was counted in 25 random fields under oil at x 1,000. The in vitro infection and the counting of infecting parasites were performed in triplicate.
Detection of host cell apoptosis
The DNA of AGS cells cultured for 24 hr and 48 hr in different pH conditions from 5.6 to 7.5 was extracted to examine apoptotic DNA fragmentation which might be induced by acidic pH. DNA extraction was done using DNeasy Tissue Kit (Qiagen Inc., Valencia, CA, USA) and followed the instruction suggested by the company. Two microgram of DNA from each sample was electrophoresed on a 1.5% agarose gel in TBE (89 mM Tris base, 89 mM boric acid, 2 mM EDTA). The gels were photographed using Polaroid Type 667 film after ethidium bromide staining.
TEM observation
The developmental stages of C. muris were observed by transmission electron microscopy (TEM). Cultured cells were washed with 0.2 M PBS (pH 7.4), and fixed in 2.5% glutaraldehyde at 4℃ for 24 hr. After washing with PBS, the cells were post-fixed with 1% osmium tetroxide for 2 hr, and then dehydrated in a graded series of ethanol and embedded in Epon. Electron microscopic observations were performed using a Jeol 1200 EXII after staining with uranyl acetate and lead citrate.
Statistical analysis
The effects of cell culture substrates, medium pH and medium supplements on C. muris infection were compared, and the statistical significance of the data was determined by ANOVA using dBSTAT™ (http://www.dbstat.com). Differences between the groups were considered statistically significant when p values were less than 0.05.
RESULTS
The number of parasites was not significantly different in cells grown on uncoated, laminin or poly-Llysine coated plates (Fig. 1A). Therefore, AGS cells were cultured onto a cover glass without further treatment with cell culture substrates. The number of infecting parasites increased with the inoculation dose (Fig. 1A), but reduced markedly with extended incubation time (Fig. 1B). Infection rates of C. muris significantly increased in lower pH culture media; the number of infecting parasites increased significantly with time passage at pH 6.6 and pH 6.2, but decreased after 24 hr of incubation at pH 7.5 (p < 0.05, Fig. 2). The incubation of AGS cells for 48 hr in medium at a pH lower than 6.4 resulted in DNA degradation of host cells, suggesting an apoptosis of host cells (Fig. 3). Therefore, subsequent studies were performed at pH 6.6 to determine the optimum conditions for in vitro cultivation, although the parasite number was highest at pH 6.2.

Effect of the surface coating of a cover glass on the in vitro development of Cryptosporidium muris in AGS cells. The number of infected C. muris was not significantly different for different coating conditions after 24 hr incubation (A), although it increased significantly (p < 0.05) as the infection dose increased. The number of infected parasites fell to less than 50% and 5% after 48 hr and 72 hr incubation, respectively, compared to that after 24 hr of incubation (B), when AGS cells were inoculated with 3 × 106 oocysts of C muris.
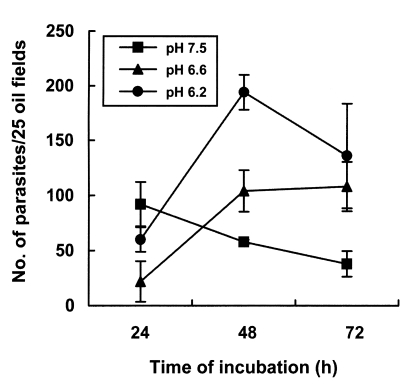
Effect of medium pH on the development of Cryptosporidium muris in AGS cells in vitro. At the beginning of the infection, pH 7.5 appeared to provide better conditions than pH 6.6 or 6.2. However, acidic pHs proved better than pH 7.5 for maintaining and continuing the in vitro infection for 72 hr.
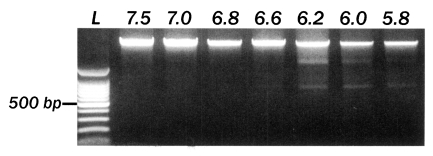
Effect of culture medium pH on the AGS cells in vitro. An incubation of AGS cells for 48 hr in medium with a pH lower than 6.2 caused DNA degradation. L, 100-bp DNA ladder. The numbers on top of each lane represent medium pH.
When cultured in three different concentrations of serum, C. muris culture in RPMI 1640 with 5% FBS resulted in 4-7 fold increases in parasite numbers versus that in 1% and 10% serum (Fig. 4A). Of the 6 medium supplements examined, 1 mM pyruvic acid was found to enhance the in vitro development of C. muris significantly (p < 0.05, Fig. 4B), whereas folic acid (2 µg/ml) produced a 20% increase (p > 0.05, Fig. 4C). However, the other supplements, i.e., sodium choleate, ascorbic acid, calcium pantothenate, and PABA either had no effect on the growth of C. muris, or significantly inhibited the parasite proliferation (p < 0.05, Fig. 4D-G).
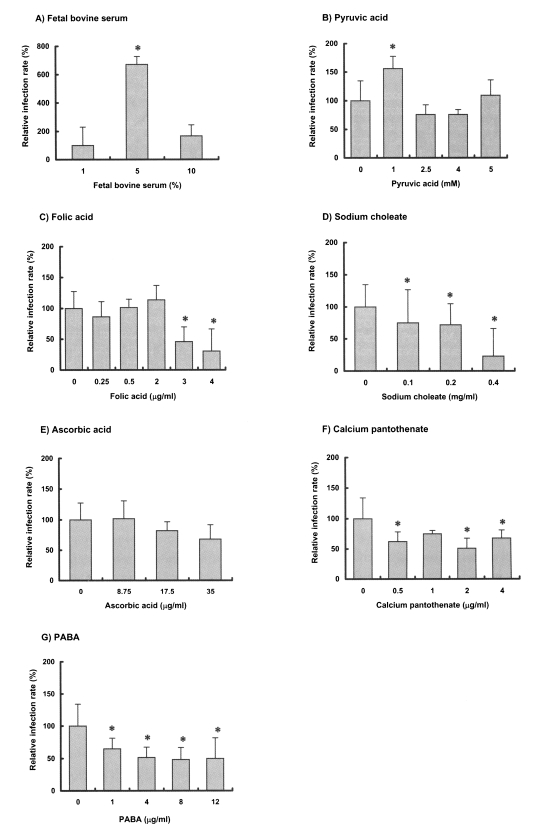
Effects of select medium supplements on the development of Cryptosporidium muris in AGS cells in vitro. Asterisks (*) denote the statistical significance of the data, compared to that of control in each medium supplement (p < 0.05).
An ultrastructural study showed the various developmental stages of C. muris in the cytoplasm of the AGS cells, which is not the typical extracytoplasmic location usually seen in the animal stomach epithelium (Fig. 5). The parasitophorous vacuolar membrane was well-formed, and separated the parasite from the host cell cytoplasm. There was a well-developed structure around attachment area between the parasite and host cell cytoplasm as filamentous cytoplasm in in vivo infection (Fig. 5A). However, the morphology of the filamentous cytoplasm-like structure observed in vitro was slightly different from that observed in vivo. In addition, the feeder organelle was difficult to find in any developmental stage. Immature oocysts were observed after 24 hr of incubation, and numerous amylopectin-like granules were found in their cytoplasm (Fig. 5B). When the cells were cultured for 48 hr, mature oocysts containing sporozoites were observed (Fig. 5C).
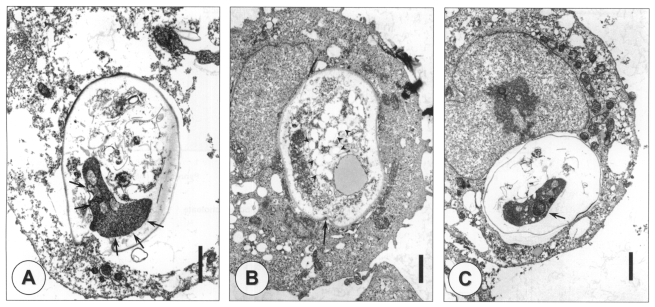
Transmission electron microscopic observation of different developmental stages of Cryptosporidium muris in AGS cells in vitro. An unidentified stage showing a filamentous cytoplasm-like structure (arrows) (A) and immature oocysts were observed at 24 hr post-infection (B), and mature oocysts containing sporozoites (arrow) were observed at 48 hr post-infection (C). Amylopectin-like granules (arrowheads) and an annular ring (arrow) are shown in an immature oocyst (B). Bar = 1 µm.
DISCUSSION
In the present study, we used AGS cells as the host cells for the in vitro culture of C. muris, because they were more susceptible to C. muris infection than Madin-Darby canine kidney (MDCK) or HCT-8 cell lines (data not shown). Considering the natural habitat of C. muris in the host, it is not surprising that AGS cells support the growth of C. muris better than MDCK or HCT-8 cells, because the AGS cell line originates from human stomach adenocarcinoma, and has an epithelial nature. The AGS cell line has also been found suitable for the in vitro culture of C. parvum (Yu et al., 2000). On the other hand, C. andersoni, which was previously referred to as C. muris in cattle, has been demonstrated to develop completely through its life cycle in HCT-8 cells in vitro (Hijjawi et al., 2002), although the abomasums, which has an acidic environment, is the normal habitat of this protozoa in cattle. However, C. muris did not grow better in HCT-8 cells than in AGS cells.
The stability of the host cell monolayer is suggested to be important for the in vitro culture of Cryptosporidium (Hijjawi et al., 2001). Moreover, sloughing of the host cell monolayer may impair the development of Cryptosporidium, and hence results in low yields of parasite in vitro. Cell cultures grown on laminin-coated plates consistently show low levels of cell detachment and an enhanced C. parvum survival in monolayers, suggesting that cell anchorage has a beneficial effect on coccidial development (Widmer et al., 2000). However, no significant increase in parasite number was observed by coating the cover glasses with cell culture substrates in this study, suggesting that the stronger attachment of AGS cell has little effect on C. muris culture.
An optimum pH appears to be important for both cell invasion and the release of the parasite stages of Cryptosporidium (Upton et al., 1995; Meloni and Thompson, 1996). A pH range of 7 to 8 resulted in the enhanced binding of C. parvum sporozoites to host cells (Hamer et al., 1994), and a neutral pH (pH 7.6) was found to be better for the excystation of C. parvum oocysts than an acidic pH (Fayer and Leek, 1984). In the present study, the number of infected C. muris increased in acidic pH's during cultivation for 72 hr, but it did not increase at pH 7.5. However, C. muris infection was thought to be inappropriate under pH 6.4 because apoptosis of the host cells occurred at a pH below 6.4. Therefore, it is suggested that an acidic pH of between 6.4 and 7.0, may be optimal for the in vitro culture of C. muris in AGS cells. Differences in the optimal pH conditions required for the in vitro cultivation of two mammalian Cryptosporidium species seem to reflect their different infection habitats in the animal host; C. muris in the stomach and C. parvum in the small intestine of their hosts.
Supplementing media with FBS or serum proteins are known to have a positive effect on the motility and development of some coccidia in vitro (Upton and Tilley, 1992). Albumin and fetuin, important serum components, enhance the motility of sporozoites and produced high numbers of the rat coccidian Eimeria nieschulzi when cultured in DMEM with 5% FBS (Upton and Tilley, 1992). In our study, the number of C. muris was greatly increased to 6.6 folds, when the medium was supplemented with 5% FBS. In the case of the in vitro culture of C. parvum, the optimal concentration of FBS has been found to be lie somewhere between 5 and 10% (Upton et al., 1995).
Of the four vitamin supplements examined, namely, L-ascorbic acid, calcium pantothenate, folic acid and PABA, folic acid at 2 µg/ml enhanced the number of C. muris oocysts slightly, but insignificantly. In contrast, the growth of C. parvum in HCT-8 cells was greatly enhanced by vitamin supplements. Ascorbic acid yielded the most pronounced effect, suggesting that ascorbic acid, an important antioxidant, enhances parasite survival (Upton et al., 1995). Therefore, vitamins appear to have less or even no effect on the development of C. muris, in contrast to their effect on C. parvum. It is difficult to explain differences in the importance of vitamins to C. parvum and C. muris culture in vitro by the present study, and needs further study.
Pyruvic acid is also known to enhance the development of some eimerians in cell culture, perhaps by channeling pyruvic acid into the TCA cycle. In the present study, the addition of 1 mM pyruvic acid resulted in a 50% increase in the number of C. muris, suggesting that pyruvic acid may provide an energy source for C. muris. Since pyruvic acid was found to be ineffective as a medium supplement for C. parvum (Upton et al., 1995), the difference between these two Cryptosporidium spp. in terms of their needs for pyruvic acid may be related with the presence of mitochondria, as C. muris has mitochondria (Uni et al., 1987), whereas C. parvum does not.
It has been reported that bile salts may affect coccidian motility, and that they induce a rapid burst of activity during excystation (Upton and Tilley, 1992). However, C. muris does not appear to need bile salts either during or following excystation. C. muris inhabits the epithelium of the stomach of mice, and therefore, there is no possibility of their encountering bile salts during their parasitism. In this study, sodium choleate showed a negative effect on the growth of C. muris in vitro; the parasite number decreased in a dose-dependent manner.
Hijjawi et al. (2001) first described the long-term maintenance of C. parvum in HCT-8 cells, and suggested the presence of an auto-reinfection cycle in cell culture. They subcultured cells every 2-3 days to remove older cells and cell debris, and maintained the parasite in vitro for up to 25 days. Thin-walled oocysts are thought to be essential for the autoinfection cycle, by facilitating the release of new invasive sporozoites from their excystation. As we observed in the present study, the presence of thin-walled oocysts is not certain.
By ultrastructural study, fully matured oocysts were observed after 48 hr of incubation, suggesting that the whole life cycle of C. muris is completed within 48 hr in the AGS cell culture system. Considering that it usually takes 2 weeks after infecting mice to detect C. muris oocysts in mice feces, the AGS cell culture allows much faster results. C. muris was incorporated in the host cell cytoplasm rather than being located extracytoplasmically. Also, characteristic structures of binding site between C. muris and epithelial cells in animal tissue, i.e., filamentous cytoplasm and feeder organelle, were not the same or found in the AGS cell culture. This differential point in AGS cell culture may imply that the in vitro infection system using AGS cells may not reflect precisely responses in vivo. Furthermore, this in vitro system is not appropriate for obtaining oocysts on a large scale. However, in terms of investigating its biological characteristics, for testing its response to therapeutic agents and for the studies on the differences between the two mammalian Cryptosporidium species, this system could provide a simple cheap alternative. The effect of medium supplements on multiplication of C. muris in vitro needs to be further evaluated in detail whether it produce the facilitative outcome either by directly on the parasite itself or by indirectly through host cells.
Notes
This study was financially supported by Konkuk University in 2001.