Intestinal histopathology and in situ postures of Gymnophalloides seoi in experimentally infected mice
Article information
Abstract
The intestinal histopathology and in situ postures of Gymnophalloides seoi (Digenea: Gymnophallidae) were studied using C3H/HeN and C57BL/6 mice as experimental hosts; the effects of immunosuppression were also observed. The metacercariae isolated from naturally infected oysters, 300 or 1,000 in number, were infected orally to each mouse, and the mice were killed at days 3-21 post-infection (PI). In immunocompetent (IC) mice, only a small number of flukes were found in the mucosa of the duodenum and jejunum during days 3-7 PI, with their large oral suckers pinching and sucking the root of villi. The intestinal mucosa showed mild villous atrophy, crypt hyperplasia, and inflammations in the villous stroma and crypt, with remarkable goblet cell hyperplasia. These mucosal changes were almost restored after days 14-21 PI. In immunosuppressed (IS) mice, displacement as well as complete loss of villi adjacent to the flukes was frequently encountered, otherwise the histopathology was generally mild, with minimal goblet cell hyperplasia. In these mice, numerous flukes were found, and it seemed that they were actively moving and rotating in situ. Several flukes were found to have invaded into the submucosa, almost facing the serosa. These results indicate that in IC mice the intestinal histopathology caused by G. seoi is generally mild, and the flukes do not penetrate beyond the mucosa, however, in IS mice, the flukes can cause severe destruction of neighboring villi, and some of them invade into the submucosa.
INTRODUCTION
After the first discovery of Gymnophalloides seoi (Digenea: Gymnophallidae) as a new human intestinal trematode in the Republic of Korea (Lee et al., 1993), medical attention has been increasingly paid to this minute fluke. One of the reasons is that a highly endemic area with 49% prevalence of the inhabitants was discovered on a southwestern coastal island (Aphae-do) of Shinan-gun, Chollanamdo (Lee et al., 1994). New endemic areas have been added (Lee et al., 1996; Sohn et al., 1998; Chai et al., 1998a, 2001). Another reason is possibility of eliciting severe intestinal pathology and symptoms by this fluke. For example, the first human case suffered from acute pancreatitis with gastrointestinal troubles (Lee et al., 1993), and two human infections were found accompanied by diabetes mellitus (Lee et al., 1995a). Many of the infected people in Shinan-gun stated that they had experienced gastrointestinal symptoms ranging from mild indigestion to severe colicky pain, although they were mixed-infected with other intestinal flukes (Lee et al., 1994). In order to understand its pathogenicity and clinical significance, the intestinal pathology caused by G. seoi, and in situ postures and behaviors of the flukes should be elucidated.
For studying the intestinal histopathology in G. seoi infection, lack of proper animal models has been a big obstacle. A natural definitive host other than man has been reported to be the palearctic oystercatcher Haematopus ostralegus, a wild bird caught in the endemic area (Ryang et al., 2000). Three other species of wild birds were experimentally proven to be highly susceptible to this fluke (Ryang et al., 2001). However, catching wild birds and keeping them in the laboratory is inconvenient and not feasible. Meanwhile, laboratory animals including gerbils, hamsters, cats, and several strains of mice (KK, C3H/HeN, ICR, BALB/c, and ddY) were found fairly susceptible to G. seoi infection, and immuno-suppression of C3H/HeN mice remarkably enhanced the worm recovery (Lee et al., 1997). The growth and development of the fluke was fairly good in this mouse strain (Chai et al., 1999). Meanwhile, in our preliminary experiments, C57BL/6 mice were found as useful as C3H/HeN mice for experimental infection with G. seoi.
The present study was performed to investigate the intestinal histopathology and in situ postures and behaviors of G. seoi in C3H/HeN and C57BL/6 mice; the effects of immunosuppression were also observed.
MATERIALS AND METHODS
Host animals and experimental groups
Specific pathogen-free mice (C3H/HeN and C57BL/6), 2~4-week-old males, 60 and 20 in each number, were purchased from the Experimental Animal Quarter, Seoul National University (Seoul, Korea). They were kept under clean conditions during the course of experiment. Three experimental groups were prepared for C3H/HeN mice; uninfected controls, immunocompetent (IC) G. seoi-infected mice, and immunosuppressed (IS) G. seoi-infected mice. The C57BL/6 mice were used for observation of the in situ postures and behaviors of the flukes; all were immunosuppressed in order to enhance survival of the flukes in their intestines (Lee et al., 1997; Chai et al., 1999). Immuno-suppression was done by intramuscular injection with a daily dose of 10 mg/kg prednisolone (Hisolong, Youngjin Pharm. Co., Korea) per mouse from day 10 prior to infection until sacrifice.
Collection of metacercariae and infection to mice
The metacercariae of G. seoi were collected from naturally infected oysters Crassostrea gigas, which were collected from the known endemic area in Shinan-gun, Chollanam-do (Lee et al., 1995b). They were isolated using the Baermann's apparatus (Beaver et al., 1984), and infected orally to mice each with 300 (C3H/HeN mice) or 1,000 (C57BL/6 mice) metacercariae using a gavage needle.
Histopathological studies of the small intestine
At days 0 (uninfected control), 3, 5, 7, 14 and 21 post-infection (PI) with G. seoi, the IC and IS mice, 5 for each group, were killed by cervical dislocation on each day. Directly after then their abdomen was opened, and 1-2 cm pieces of their small intestines (duodenum, proximal jejunum, middle jejunum, distal jejunum, and ileum) were quickly resected, followed by fixation in 10% neutral formalin. The fixed tissues were dehydrated in graded series of alcohol, embedded in paraffin, cut into 5-7 µm sections, and stained with hematoxylin and eosin.
RESULTS
Immunocompetent mice
Uninfected control mice (C3H/HeN) revealed their normal contour of the mucosa of the small intestine, with tall, slender villi, and villus/crypt (V/C) height ratio of 4-5, except for slight non-specific inflammations in the villous stroma (Fig. 1). On the other hand, the intestines of G. seoi-infected (with 300 metacercariae) IC mice (C3H/HeN) typically showed villous atrophy (V/C ratio: 3) and crypt hyperplasia in the duodenum and jejunum, with a moderate degree of inflammatory reactions in the villous stroma and crypt (Figs. 2-8). Despite such mild pathological changes, goblet cell hyperplasia was remarkable along the villous epithelia (Figs. 2, 3). The goblet cell hyperplasia was more extensive in the jejunum (Figs. 2, 3) than in the duodenum (Fig. 4) or ileum. At day 5 PI, adult flukes of G. seoi were characteristically seen pinching and sucking the epithelial layer of villi with their oral suckers (Figs. 3, 4). Appearance of the flukes in intestinal sections was not so frequent.
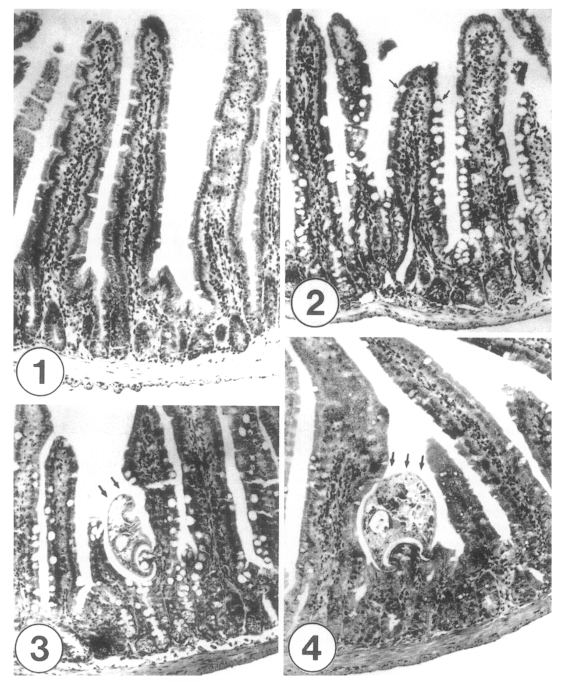
Sections of the small intestines of a normal control C3H mouse (Fig. 1) and those infected with 300 metacercariae of Gymnophalloides seoi, day 5 post-infection (PI) (Figs. 2-4). × 100. Fig. 1. Duodenum of a control mouse showing their normal tall and slender villi with non-specific stromal inflammation, and normal crypts. Fig. 2. Proximal jejunum of a mouse infected with G. seoi. Note the remarkable goblet cell (arrows) hyperplasia along the epithelial layer of the villi, inflammatory reactions in the crypts, and increased secretory granules in paneth cells of the crypts. Fig. 3. Middle jejunum of a mouse infected with G. seoi. An adult fluke (arrows) is characteristically pinching the epithelial layer of a villus with its oral sucker. Goblet cell hyperplasia and cryptitis are remarkable. Fig. 4. Duodenum of a mouse infected with G. seoi. An adult fluke (arrows) is seen pinching the root portion of a villus with its large oral sucker. Crypts show inflammatory reactions.
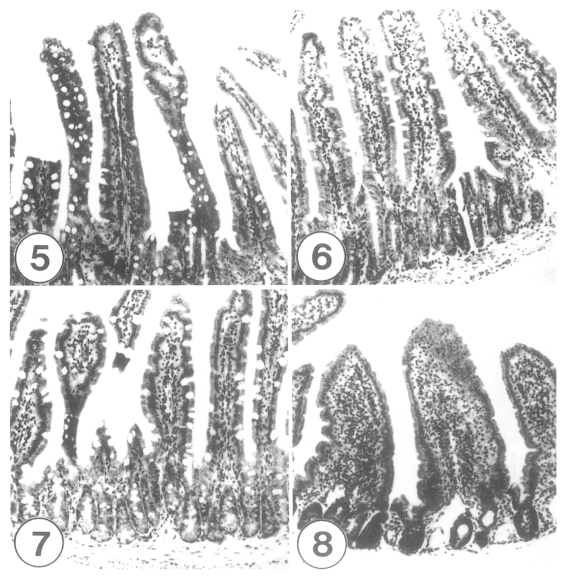
Sections of the small intestines of C3H mice infected with 300 metacercariae of Gymnophalloides seoi, days 14-21 post-infection (PI). × 100. Fig. 5. Middle jejunum of a mouse at day 14 PI, showing tall and slender villi, but with stromal inflammation and goblet cell hyperplasia. Fig. 6. Duodenum of a mouse at day 21 PI. Note that the shape of the villi is normal, and goblet cell hyperplasia is not recognizable in this portion of the small intestine. Fig. 7. Middle jejunum of a mouse, day 21 PI. Slight goblet cell hyperplasia and crypt hyperplasia are still recognizable. Fig. 8. Proximal jejunum of a mouse, day 21 PI. Blunting, fusion, and edema of villi with stromal inflammations are prominent.
These pathological changes were almost restored at days 14-21 PI (Figs. 5-8). The mucosa showed tall and slender villi (V/C ratio: 4), although stromal inflammation and goblet cell hyperplasia sustained, especially in the jejunum (Figs. 5, 7). In the duodenum, goblet cell hyperplasia was no more recognizable (Fig. 6). However, crypt hyperplasia as well as villous atrophy including blunting, fusion, and edema of villi, with stromal inflammations remained focally in the jejunum at day 21 PI (Figs. 7, 8).
Immunosuppressed mice
Uninfected control mice (C3H/HeN) immunosuppressed with prednisolone revealed nearly normal contour of the small intestinal mucosa, with slightly decreased V/C ratio (3.5-4.5), and decreased intestinal wall thickness. In G. seoi-infected (300 metacercariae) IS mice (C3H/HeN), the histopathological features (Figs. 9-16) were similar to those observed in G. seoi-infected IC mice. The destruction of villi adjacent to the flukes was, however, generally severer than in IC mice. At days 3-7 PI, inflammatory reactions and vascular congestion were frequently seen in the villous stroma and the crypt (Fig. 9). The villous atrophy and crypt hyperplasia were moderate or severe (Fig. 10). Many juvenile or adult flukes were seen occupying the intervillous spaces (Figs. 9-12). They were pinching (and looked-like eating) the root of villi with their large oral suckers (Figs. 9, 10, 12). Around the flukes, pressure-atrophy of the villous epithelia and at times complete loss of villi were encountered (Figs. 10-12). Mild goblet cell hyperplasia was recognized in some sections (Fig. 13), but generally not remarkable in IS mice (Figs. 9-12).
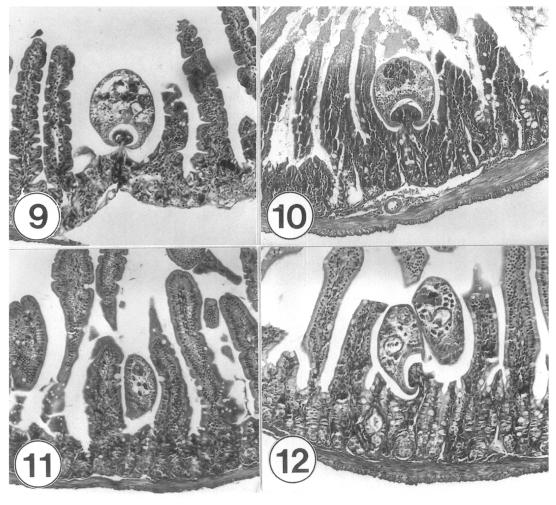
Sections of the small intestines of immunosuppressed C3H mice experimentally infected with 300 metacercariae of Gymnophalloides seoi, days 3-7 post-infection (PI). × 100. Fig. 9. Proximal jejunum of a mouse infected with G. seoi, at day 3 PI. A juvenile fluke is seen pinching (looks like eating) the root portion of a villus with its large oral sucker. The villous stroma and the crypt show congestion and inflammatory reactions. Fig. 10. Proximal jejunum of another infected mouse, at day 3 PI. A juvenile fluke is characteristically seen pinching the root portion of a villus with its oral sucker. Fig. 11. Proximal jejunum of another infected mouse, day 5 PI. Note the pressure-atrophied epithelium of villi adjacent to a fluke. Goblet cell hyperplasia is not remarkable. Fig. 12. Proximal jejunum of another infected mouse, day 7 PI. Two adult flukes, with uterine eggs, are seen to have caused pressure atrophy of the surrounding epithelia of the villi. One of the flukes is pinching the root of an adjacent villus.

Sections of the small intestines of immunosuppressed C3H mice experimentally infected with 300 metacercariae of Gymnophalloides seoi, days 7-21 post-infection (PI). × 100. Fig. 13. Ileum of an infected mouse, day 7 PI. A low-grade goblet cell hyperplasia is seen on the epithelial layers of the villi and crypts, but the contour of the villi was normal. Crypts show inflammatory reactions. Fig. 14. Proximal jejunum of an infected mouse at day 14 PI. Note that the shape of the villi is normal, and goblet cell hyperplasia is not recognizable, although inflammation remained in the crypts. Fig. 15. Proximal jejunum of an infected mouse, day 21 PI. Despite the presence of a fluke in the lumen, the contour of the villi are normal, except inflammations in the villous stroma and inflammation and dilatation of the crypts. Goblet cell hyperplasia is no more recognizable. Fig. 16. Ileum of an infected mouse, day 21 PI. Villi show their normal contour, but crypt hyperplasia, with stromal inflammation, is seen. × 100.
At days 14-21 PI, the shape of the villi became almost restored, and goblet cell hyperplasia was no more recognizable (Figs. 14-16). However, inflammation in the villous stroma and crypts remained over the small intestine (Figs. 14-16).
In situ postures of flukes
In IS mice (C57BL/6) infected with G. seoi (1,000 metacercariae), numerous adult flukes were found pinching and sucking the root of the host intestinal villi (Figs. 17, 18), from which behaviors of the flukes could be imagined. Almost all flukes were containing a fragment of host intestinal tissue in their oral suckers. Displacement as well as complete loss of the infected villi was seen nearby the groups of flukes.
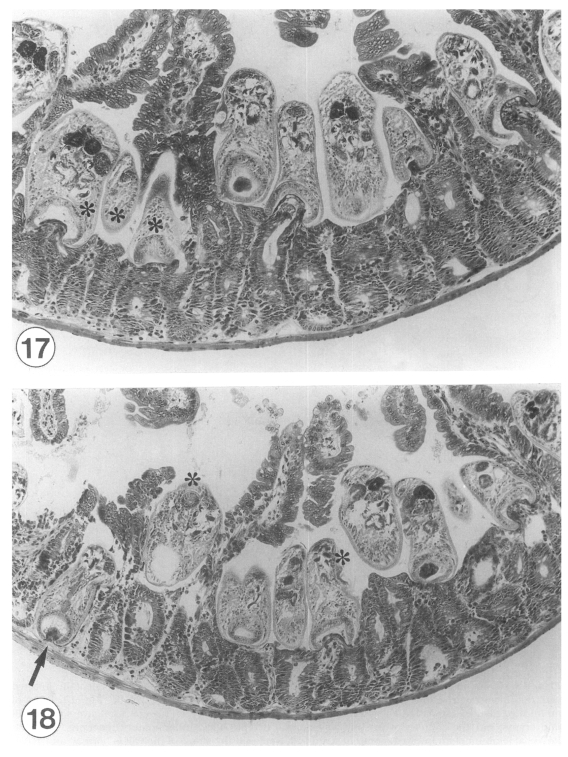
Sections of Gymnophalloides seoi adults in the small intestine of an immunosuppressed C57BL/6 mouse at day 7 post-infection (PI) with 1,000 metacercariae. × 250. Fig. 17. In situ postures of adult flukes in the middle jejunum of an infected mouse, from which their behaviors could also be imagined. The flukes are pinching and sucking the root portion of the host villi, and displacement as well imagined. The flukes are pinching and sucking the root portion of the host villi, and displacement as well as loss of the affected villi are seen nearby a group of worms (*). Fig. 18. Adult flukes attached to the mucosa of the jejunum of an infected mouse. A worm (arrow) is seen to have penetrated deeply into the submucosa, just facing the serosa, and keeping a fragment of mucosal tissue within its oral sucker. Many flukes contained eggs in their uteri (*), and showed other organs such as the oral and ventral suckers, vitellaria, intestinal ceca, pharynx, testes, and seminal vesicle.
A peculiar finding was that a few adult flukes were seen to have penetrated deeply into the submucosa, just facing the serosa (Fig. 18). These flukes were also keeping a fragment of mucosal tissue within their oral suckers (Fig. 18). Many flukes contained eggs in their uteri, and reproductive and parenchymatous organs were conspicuous.
DISCUSSION
With regard to the host intestinal histopathology due to intestinal fluke infections, many studied have documented villous atrophy and crypt hyperplasia with inflammatory reactions in the villous stroma and crypts as the most important findings (Chai and Lee, 1990). The species of flukes subjected for studies include Metagonimus yokogawai (Chai, 1977; Lee et al., 1981; Kang et al., 1983; Chai et al., 1994a, 1995), Neodiplostomum seoulense (Lee et al., 1985), Pygidiopsis summa (Seo et al., 1986), and Echinostoma hortense (Lee et al., 1990). The results of the present study for G. seoi were similar to those reported for other species of intestinal flukes, and they seem to be common histopathological features for intestinal flukes general. However, in the present study, the intestinal histopathology was relatively mild compared to other species of flukes except for strong goblet cell hyperplasia in G. seoi-infected IC mice (C3H/HeN).
One of the reasons for the mild histopathology seems to be the small number of flukes actually parasitizing in the small intestine of IC mice. For example, the worm recovery of G. seoi from IC C3H/HeN mice was reported to be about 12% at day 7 PI (Lee et al., 1997). Therefore, when 300 metacercariae were given to each mouse, about 36 adult flukes would have survived in the small intestines of these mice. Such a small number of flukes seem not sufficient to cause severe histopathological changes. Another reason may be relatively low pathogenicity of G. seoi compared to other intestinal flukes like echinostomes, which caused severe pathology with ulceration of the mucosa and bleeding from the infected sites (Lee et al., 1990; Chai et al., 1994b). Cysteine proteinases were purified from metacercariae and adults of G. seoi, followed by functional characterizations (Choi et al., 1998a, 1998b). These enzymes were suggested to be involved in nutrient uptake and immune evasion from the host, but not in eliciting pathogenicity to the host.
Meanwhile, mechanical irritation by the flukes is considered to be the major possible mechanism for the pathogenicity by G. seoi. The immunosuppression increased significantly survival of the flukes in IS mice than in IC mice, and in IS mice, villi adjacent to the flukes were severely destroyed. In some sections, many adult flukes were pinching and sucking the root of villi, with their large oral suckers. Villi nearby the flukes were pressure-atrophied, and some areas showed complete loss of villi. Therefore, it is suggested that immunosuppression enhanced survival of many flukes in the host intestinal mucosa, then the flukes should have caused mechanical irritation on the neighboring villi and crypts. Probably owing to immuno-suppression, however, host systemic immune responses must have been compromised, and mucosal inflammation and pathology other than destruction of villi appeared to be generally mild.
A striking observation among the histopathological features in the small intestines of G. seoi-infected IC mice was strong proliferation of goblet cells on the villous epithelia. This feature is of considerable significance to understand the host-parasite relationships in G. seoi infection, since goblet cells were reported as an important effector for natural expulsion of intestinal helminths; e.g., Nippostrongylus brasiliensis (Nawa et al., 1994; Onah and Nawa, 2000) and Echinostoma trivolvis (Fujino et al., 1993). However, mucosal immune mechanisms for the expulsion of intestinal helminths have been known to be rather complex. Mucosal mast cells but not goblet cells were shown to be the important effector for the expulsion of Strongyloides ratti (Abe et al., 1992, 1993). On the other hand, goblet cells and mast cells played only minor roles in host defenses against Neodiplostomum seoulense infection (Chai et al., 1998b).
Exact immune effector mechanisms implicated in the expulsion of G. seoi from the mouse intestine have not been documented until present. However, it is strongly suggested by the present study that goblet cells in the mucosa would be one of the important effectors responsible for the short-time parasitism of G. seoi in rodent hosts (Lee et al., 1997; Chai et al., 1999). This suggestion is supported by the strong goblet cell responses in IC mice during days 3-7 PI, and only a small number of flukes retained in their intestines at days 7-14 PI. It is further supported by the minimal degree of goblet cell hyperplasia in IS mice, with much more flukes surviving in the intestine. The detailed mechanisms of goblet cell involvement in the expulsion of G. seoi, however, remain to be elucidated.
Difficulties were met for observation of the in situ postures of G. seoi in IC and IS C3H/HeN mice, both of which were given 300 metacercariae per mouse. More flukes were found sectioned in IS mice than in IC mice, however, not yet satisfactory. Therefore, to obtain a sufficient number of worm sections, the C57BL/6 mice were infected with 1,000 metacercariae, and all of them were immuno-suppressed. Numerous flukes were sectioned in the intestines of these mice, which greatly helped observation of the in situ worm postures and behaviors.
The flukes were found in situ pinching, sucking, and eating the root portion of the intestinal villi with their big oral suckers. They were frequently seen keeping fragments of mucosal tissues within the oral sucker. These postures were essentially the same as those observed in their second intermediate host, being attached to the mantle surface of the oysters (Lee et al., 1995b). Other behaviors of the flukes in the mouse intestine seem to include vigorous moving and rotating in situ and giving pressures to the neighboring villi. Eventually, displacement as well as complete loss of villi has occurred. In IC C3H/HeN mice, the flukes were never found beyond the mucosa of the small intestine. By contrast, in IS C57BL/6 mice, some of the flukes were found in the submucosa and deeper layers, almost facing the serosa.
It is worth while to note that the first patient of G. seoi suffered from acute pancreatitis (Lee et al., 1993), and two other cases accompanied diabetes mellitus (Lee et al., 1995a). It was thus suspected that G. seoi could infect the pancreatic duct in the human host. Other gymnophallids were found infected in the bursa Fabricii and gallbladder of shore birds as well as their intestines (Yamaguti, 1971; Schell, 1985). However, in our series of studies using rodent animals as the host (Lee et al., 1995b, 1997; Chai et al., 1999), G. seoi have never been found from locations other than the intestinal tract, which is difficult to understand. Bigger experimental animals may be needed to verify the possibility for G. seoi to infect the pancreatic duct or other locations. Moreover, as the flukes invaded into the submucosa of IS mice in the present study, the possibility for G. seoi eggs to be transferred to remote organs in immunocompromised hosts should be ruled out.
The results of the present study suggest that the pathogenicity of G. seoi may not be so severe in immunologically normal hosts. However, in immunocompromised hosts, G. seoi may invade into the submucosa and other locations.
Notes
This work was supported in part by Basic Medical Science Research Fund, Ministry of Education, Republic of Korea (1997).