Chromosomes of the liver fluke, Clonorchis sinensis
Article information
Abstract
A karyological study was carried out in order to compared the chromosome numbers, chromosome morphologies and karyotypes of the oriental liver fluke, Clonorchis sinensis (Trematoda: Opisthorchiidae), collected from Korea and China. Chromosome preparations were made by means of air-drying method. The chromosome number was 2n=56 in both Korean and Chinese flukes, and chromosomes were divided into two groups based on this size; consisting of 8 pairs of large and 20 pairs of small chromosomes. However, the karyotypes showed some differences between Korean and Chinese flukes. The karyotype of liver flukes from Korea consisted of three metacentric pairs, one meta-/submetacentric pair, 16 submetacentric pairs and eight subtelocentric pairs of chromosomes. On the other hand, liver flukes from China consisted of two metacentric pairs, two meta-/submetacentric pairs, 16 submetacentric pairs and eight subtelocentric pairs of chromosomes.
INTRODUCTION
Clonorchiasis is an important food-borne parasitic zoonosis in East Asia, being endemic in Korea, Japan, and all parts of China except the northwest, Taiwan and Vietnam. Although reports of this infection are infrequent in western countries, infection can be acquired by eating frozen, dried, or pickled freshwater fish imported from endemic areas (Rim, 1986, 1990). People in endemic areas acquire infection by eating raw or uncooked freshwater fish.
Cytotaxonomic studies of the parasites are important not only for the parasites of systematic analyses, but also for the basic information of the parasitic agents. From this point of view, chromosomal studies on trematode parasites have been noticeably useful in systematic studies (Sakakuchi and Nakagawa, 1975; Sakaguchi and Tada, 1975; Hirai et al, 1985; Mutafova and Kanev, 1996; Terasaki et al., 1996; Park et al., 1998); but, until recently there have been relatively a few studies on the chromosomes of trematode parasites. Barsiene (1993) reviewed chromosome numbers and karyological data for 230 species of trematodes. Diploid chromosome numbers of the trematodes ranged from 16-22 (Short et al., 1989; Terasaki et al., 1995). Until now, no studies on the chromosome of Clonorchis sinensis from Korea and China have been attempted.
The purpose of this study was to analyze the chromosome numbers and karyotypes of the liver fluke, C. sinensis, collected from Korea and China.
MATERIALS AND METHODS
Metacercariae were removed from a freshwater fish, Pseudorasbora parva, collected at Kimhae, Kyungsangnam-Do, Korea and at Shenyang, Liaoning Province, China. Metacercariae were orally inoculated into rabbits. Rabbits were sacrificed 3-5 months after infection and the adult flukes obtained were subjected to cytological examination. The chromosomes in the gonadal tissues of the fluke were studied by the simple cell cultivation method of Ando and Uchida (1973) with minor modification and by Park's (1994) air-drying method. Morphological features of the chromosomes used to compare karyotypes were the relative length and total lengths of the mitotic metaphase chromosomes. Relative length was expressed as the ratio of absolute chromosome pair length over the total length of the haploid complement ×100. The prepared slides were observed under an Olympus BX50 microscope with a 100X oil immersion objective and a 10X ocular. The nomenclature of the centromeric position for the classification of chromosomes was followed as proposed by Levan et al. (1964).
RESULTS
Mitotic chromosomes were observed in 118 cells from 12 individuals in Korea and in 121 cells from 18 individuals in China. All of the metaphase figures of mitosis in C. sinensis, 56(2n) chromosomes were recognized (Figs. 1-4). According to the size and shape of the chromosomes, the karyotypes were established on plates for 28 pairs. Although there were size variations in the chromosomes, the complements were tentatively ordered into pairs according to size. They could be divided into two groups by size, 8 pairs of large and 20 pairs of small chromosomes. Table 1 shows the measurements of mitotic metaphase chromosomes in three sets of karyotyped cells from Korea and five sets of karyotyped cells from China population. The karyotypes of Korean population consisted of three metacentric pairs, one meta-/submetacentric pair, 16 submetacentric pairs and eight subtelocentric pairs of chromosomes (Fig. 2). The karyotypes of Chinese population consisted of two metacentric pairs of chromosomes, two meta-/submetacentric pairs, 16 submetacentric pairs and eight subtelocentric pairs of chromosomes (Fig. 4.). The mean lengths of the mitotic metaphase chromosomes ranged from 12.01 ± 0.10 µm to 1.73 ± 0.43 µm from Korea and 11.08 ± 0.16 µm to 2.03 ± 0.15 µm from China. The mean total length of diploid complement was 136.64 ± 5.1 µm in Korea and 144.44 ± 4.39 µm in China. The shapes of the various diakinesis bivalents that have been observed in C. sinensis are shown in Figure 5 and 6.
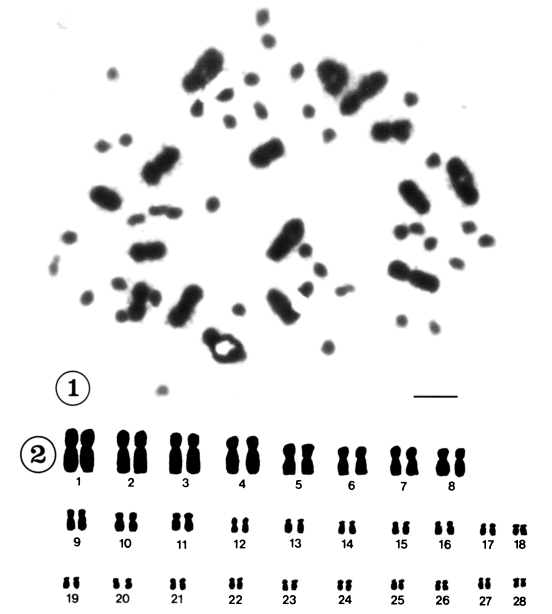
Mitotic chromosomes of Clonorchis sinensis from Korea. Fig. 1. Diploid chromosomes in metaphase I. Scale bar=10 µm. Fig. 2. Camera lucida drawings of Fig. 1.
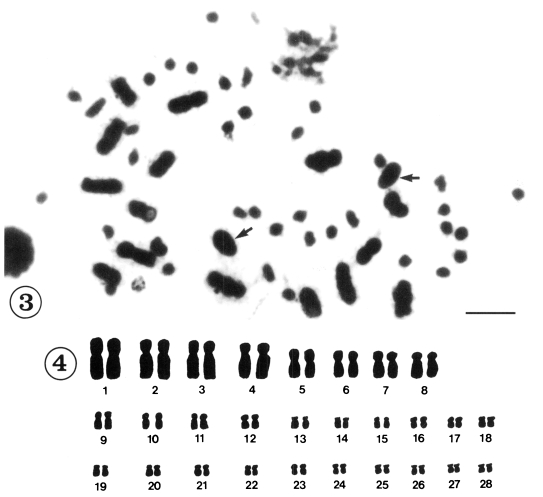
Mitotic chromosomes of Clonorchis sinensis from China. Fig. 3. Diploid chromosomes in metaphase I. Arrows indicate sperm heads. Scale bar= 10 µm. Fig. 4. Camera lucida drawings of Fig. 3.

Total lengths (μm) and relative lengths of chromosomes of Clonorchis sinensis from Korea and China
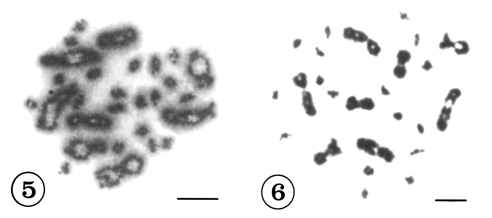
The meiosis diakinesis bivalents of Clonorchis sinensis. Fig. 5. C. sinensis from Korea. Fig. 6. C. sinensis from China. Scale bars = 10 µm
DISCUSSION
The morphology of chromosomes among individuals of the same species is sufficiently constant that karyotypes are recognized as a definite species character (White, 1973). The study of chromosome numbers has been used as a valuable complement to biochemical methods for identifying species, hybrids and more rarely populations. It has also been of particular importance in the application of polyploidy. In fact, relationships within groups of species cannot be considered complete, in an evolutionary sense, without good cytotaxonomic data to reinforce conclusions drawn from morphological criteria.
Barsiene (1993) reviewed chromosome numbers and karyological data for 230 species of trematodes. Chromosome numbers of trematodes ranged from 2n = 16 for Schistosoma mansoni (Short et al., 1989), 2n = 20 for Fasciola spp. (Yin and Ye, 1990), Neodiplostomum seoulense (Park et al., 1998), Pegosomum asperum and P. saginatum (Aleksandrova and Podgornova, 1978) to 4n = 44 for Paragonimus westermani (Terasaki et al., 1995). In this study, the chromosome number of C. sinensis from Korea and China was 2n = 56. Until now, no C. sinensis have been investigated cytologically prior to this study. Karyotype studies prove of greatest value to systematic study when used with more closely related species. The mean total length of the diploid complements of liver flukes from China was a little longer than those from Korea. Sasaki (1961) stated that individual chromosomes in the metaphase stage usually exhibited considerable variation in length due to varying degrees of condensation on decomposing chromatids on mammalian somatic chromosomes. By his result, highly contracted chromosomes tend to have their centromeres more median than less contracted ones in different cells, while longer chromosomes tend to contract more strongly than shorter ones in the same cell by colchicine treatment. In this chromosomal study, we made preparations in the same conditions. We expressed the lengths of individual chromosomes as relative lengths in terms of a percentage of the haploid autosomal complement. Actually, this air-drying technique produces a more uniform and complete flattening of cells than can be achieved by the usual squash method.
The karyotypes of C. sinensis from Korea and China were different. Differences in chromosome morphology could be due to inversions, additions, deletions, etc. which might still represent within species variability. Nonetheless, this speculation enhances the authors' conclusion that more study is warranted for these groups of flukes. This chromosomal data will provide basic information on the liver flukes, which can then be used for genome study.