Cryptosporidium parvum is a parasitic protozoan that infects gastrointestinal epithelial cells of many vertebrates, including humans [1]. It causes watery diarrhea and can be fatal to immunocompromised individuals [1]. There are 20 known Cryptosporidium species and at least 44 genotypes, which differ significantly in their molecular signatures but have not been assigned species status [2]. Eight fully characterized Cryptosporidium species (C. hominis, C. parvum, C. meleagridis, C. felis, C. canis, C. suis, C. muris, and C. andersoni) and 5 partially characterized species (from the deer, monkey, skunk, rabbit, and chipmunk) infect humans [2-5], among which C. hominis and C. parvum are the most commonly detected [3]. A real-time PCR (qPCR) method using primers derived from the CP2 gene is highly sensitive, specific, and accurate for the detection of cryptosporidiosis but cannot distinguish among species [6]. Therefore, we developed a qPCR-based restriction fragment length polymorphism (RFLP) method for the 2 Cryptosporidium spp. differentiation and we detected C. hominis in 1 of 21 patients with diarrhea.
DNA was prepared from C. parvum oocysts of (KKU isolates) collected from laboratory mice (C57BL6/J) that were infected with the parasite and maintained as described [6]. DNA was extracted from the oocysts and fecal materials by using a QIA-quick Stool Mini Kit (QIAGEN Inc., Hamburg, Germany USA). The qPCR reactions were performed according to the method of Lee et al. [6] using a LightCycler® (Roche, Basel, Switzerland, USA). The results were analyzed using the LightCycler® software (version 4.05, Roche). DNase/RNase-free water was used in place of template DNA as a negative control. CP2 sequences of C. parvum (AY471868) and C. hominis (XM_661199) were aligned using Clone Manager Suite 7 (Sci-Ed Software, Cary, North Carolina, USA), and restriction enzyme cleavage sites were identified using NEBcutter V2.0 (http://tools.neb.com/NEBcutter2/). An aliquot (15 µl) of the qPCR product was digested with TaqI (Takara Bio Inc., Shiga, Japan) at 65℃ for 2 hr, and DNA fragments were analyzed using 2.5% agarose gels. Stool samples from 21 patients with diarrhea in the Busan area of Korea were collected from June 1 to 30, 2011, by the Korea Centers for Disease Prevention and Control. Modified acid-fast staining was performed on stool samples that were positive for C. hominis by using qPCR-based RFLP.
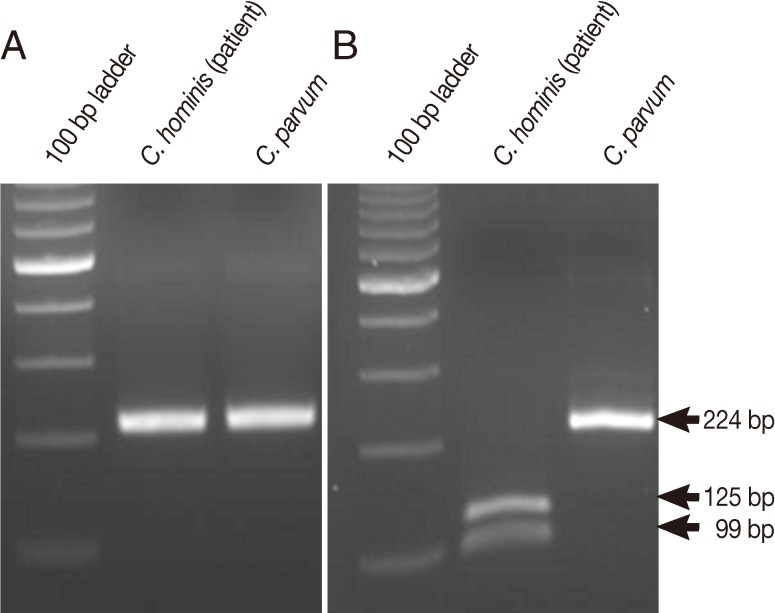
The sequences of the CP2 genes of C. parvum (GenBank AY-471868) and C. hominis (GenBank XM_661199) are 94% identical (data not shown). Because analysis using NEBcutter V2.0 identified a TaqI site only in the sequence of C. hominis
CP2 (Table 1), TaqI was chosen for restriction fragment length polymorphism (RFLP) analysis.
Using qPCR, we found that 1 of 21 patients with diarrhea tested was positive for CP2 (data not shown). The patient with positive results was a 6-year-old girl with watery diarrhea. No other information about the patient was available. To further identify the species of Cryptosporidium responsible for the infection, the sample was digested using TaqI, which generated 99 bp and 125 bp bands, indicating that the patient was infected by C. hominis (Fig. 1). C. hominis oocysts were also detected in the diarrheal stool sample by using the modified acid-fast staining (Fig. 2).
Morgan-Ryan et al. [7] proposed a new species of Cryptosporidium, C. hominis, to indicate its isolation from human feces. However, C. hominis and C. parvum oocysts are morphologically indistinguishable [7]. Species discrimination is important for molecular epidemiological purposes to evaluate potential sources of infections [8]. Real-time PCR increases the speed of sample analysis and decreases the risks of contamination with DNA present in the laboratory [8]. The present study showed that both major Cryptosporidium species can be detected simultaneously and distinguished from each other by using TaqI to digest the CP2 gene of C. hominis. Because the CP2 gene is highly specific, no genetic information is available for other Cryptosporidium species, except for C. parvum and C. hominis in GenBank (www.ncbi.nlm.nih.gov/genbank/).
The genotypes of Cryptosporidium in Korea have been reported [9-11]. Cheun et al. [11] studied Cryptosporidium sp. in 3 rural areas by using a PCR-RFLP method to detect 18S rDNA sequences and identified C. parvum in 12 patients with Cryptosporidium infection. Therefore, the case confirmed by the present study is very important, because it indicates the presence of C. hominis infection in Korea.
In the present study, we developed a simple and accurate qPCR-based RFLP method for differentiating C. parvum from C. hominis. This method could be helpful in facilitating the detection of C. hominis infection in Korea.