Surface ultrastructure of Pygidiopsis summa (Digenea: Heterophyidae) adult flukes
Article information
Abstract
A scanning electron microscopic study was performed on the surface ultrastructure of Pygidiopsis summa (Digenea: Heterophyidae) adults. Metacercariae were collected from gills and muscles of mullets (Mugil cephalus) caught in a known endemic area, and adult flukes were harvested from dogs after 8 weeks of experimental infection. The worm was calabash form with its posterior part broader than the anterior part. Tegumental spines were densely distributed over the body surface, except on the suckers and genital apparatus, and around the excretory pore. Well differentiated spines were observed on the anterior half of the body, with 14-16 tips ventrally, and 19-20 tips dorsally. On the oral sucker, three pairs of type I sensory papillae (uni-ciliated knob-like swellings) and one pair of type II sensory papillae (aciliated round-swellings) were observed on the anterior and posterior parts of the lip, respectively. On the lip of the ventral sucker, one pair of type II sensory papillae was distributed only on its posterior part. Sperms were seen emerging from or entering into the genital apparatus. The results showed that the surface ultrastructure of P. summa was unique among the heterophyid trematodes, especially in digitation of tegumental spines and in distribution of sensory papillae on oral and ventral suckers.
INTRODUCTION
Pygidiopsis summa (Digenea: Heterophyidae) was originally described in Japan from dogs experimentally fed mullets infected with the metacercariae (Onji and Nishio, 1916). Human infections were first found by detection of eggs in the feces in Japan (Takahashi, 1929), and adult flukes from humans were identified later (Yokogawa et al., 1965). This parasite is now known to distribute in the Republic of Korea (Chun, 1963; Seo et al., 1981a, 1981b; Chai and Lee, 1990; Sohn et al., 1994), and at least 31 worm-proven human cases have been reported (Seo et al., 1981a; Chai et al., 1997, 1998a). The adult flukes are morphologically characterized by their small concave body, median location of the ventral sucker, unique morphology of the ventrogenital apparatus, and side-by-side location of two testes (Chai et al., 1986).
Surface ultrastructures of helminth parasites, as revealed by scanning electron microscopy (SEM), are helpful for not only taxonomic studies but also for immunological and drug efficacy studies. In heterophyid trematodes, SEM studies have been performed in Cryptocotyle lingua (Koie, 1977), Heterophyes aequalis (Taraschewski, 1984), H. nocens (Chai et al., 1992), Heterophyopsis continua (Hong et al., 1991), Metagonimus yokogawai (Lee et al., 1984), M. miyatai (Chai et al., 1998b), and M. takahashii (Chai et al., 2000). However, no information is available for P. summa. Thus, the present study was performed to observe the surface ultrastructure of P. summa adults and to compare the results with those reported for other heterophyid trematodes.
MATERIALS AND METHODS
Metacercariae of P. summa were isolated from gills and muscles of the mullet (Mugil cephalus) caught at an estuary in Okku-gun, Jeonrabuk-do (Province), by artificial digestion technique. Adult flukes were harvested from the small intestine of experimentally infected dogs at week 8 post-infection (PI). The adult flukes were washed with 0.2M cacodylate buffer (pH 7.3) and fixed in 2.5% glutaraldehyde. They were dehydrated, dried, and mounted on aluminum stubs, followed by coat with gold. The specimens were observed with a scanning electron microscope (ISI DS-130C, Korea) at an accelerating voltage of 10 kV.
RESULTS
Adult flukes are ventrally concave, calabash or pyriform in shape, and equipped with oral, ventral suckers, and a genital apparatus (Fig. 1). The whole body surface is covered with flat cytoplasmic processes, numerous tegumental spines, and sensory papillae, as seen from ventral (Figs. 1-7) and dorsal views (Figs. 8-14).
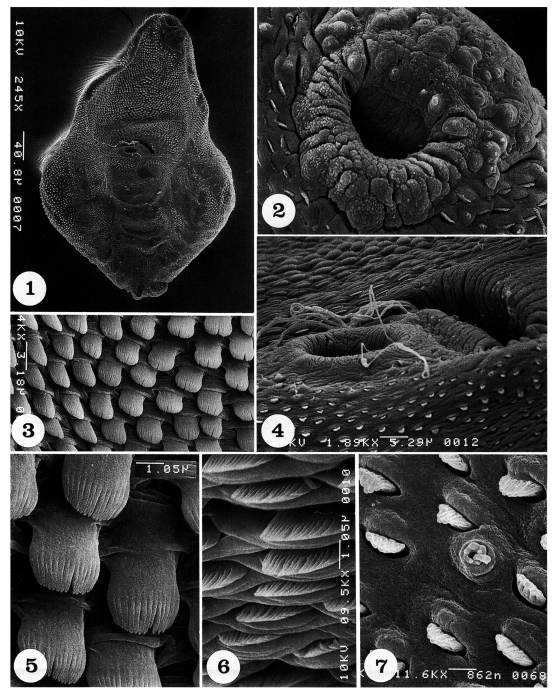
Scanning electron micrographs of Pygidiopsis summa adults (ventral view). Fig. 1. A whole adult fluke, showing its calabash form with oral and ventral suckers and the genital apparatus. Fig. 2. Tegument around the oral sucker. Note the distribution of one pair of small type I sensory papillae (arrowheads) and two pairs of large type I sensory papillae on the anterior part of the lip of the oral sucker. On the posterior part of the lip, a pair of type II sensory papillae are seen medially. Fig. 3. Tegument between oral and ventral suckers, showing dense distribution of tegumental spines, which are long painting brush-shape and have 14-16 digits. Fig. 4. Tegument around the ventral sucker and genital apparatus. Note that sperms are escaping from or entering into the genital pore. Fig. 5. Magnification of the tegumental spines in Fig. 3. Fig. 6. Tegument just anterior to the genital apparatus, showing short broom brush-shaped tegumental spines, having 14-16 digits. Fig. 7. Tegument on the posterior body surface, showing a uni-ciliated type I sensory papilla and several spade-shaped tegumental spines with 5-8 digits.
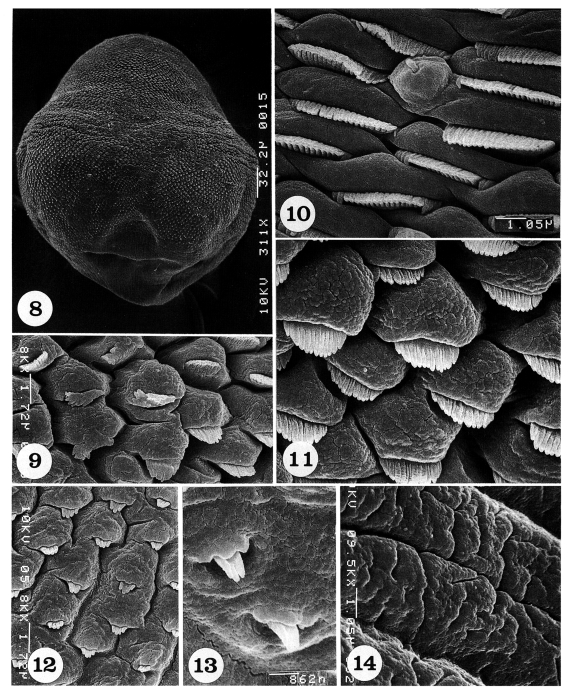
Scanning electron micrographs of Pygidiopsis summa adults (dorsal view). Fig. 8. A whole adult fluke, showing numerous tegumental spines on the body surface, except around the posterior end of the body. Fig. 9. Tegument on the antero-lateral body surface, showing protruded cytoplasmic processes, each with a short tegumental spine having 15-17 digits. Fig. 10. Tegument on the mid-lateral body surface, showing a uni-ciliated sensory papilla and short wide tegumental spines with 19-21 digits. Fig. 11. Tegument on another mid-lateral body surface, showing protruded cytoplasmic processes, each with a spade-shaped tegumental spine having 12-14 digits. Fig. 12. Tegument on the mid-posterior body surface, showing protruded cytoplasmic processes, each with a bear claw-shaped tegumental spine having 4-5 digits. Fig. 13. Tegument on the postero-lateral body surface. Spines are sparsely distributed and claw-shaped with 3-4 digits. Fig. 14. Tegument near the posterior end of the body, wrinkled and devoid of spines.
The oral sucker is small and located near the anterior end (Fig. 2). It has one pair of small type I sensory papillae (ciliated knob-like swellings) and two pairs of large type I sensory papillae on the anterior part of the lip, and one pair of type II papillae (aciliated round-swellings) on its posterior lip (Fig. 2). On the tegument between oral and ventral suckers, spines are densely distributed; they are long, painting brush-shaped, and having 14-16 digits (Figs. 3 & 5). The ventral sucker is small, submedian, protruded ventrally, and equipped with one pair of type II sensory papillae on its postero-lateral margins (Fig. 4). Just anterior to the ventral sucker, the genital apparatus is seen as a transversely long furrow, and sperms are seen escaping from or entering into the genital pore (Fig. 4). On the tegument anterior to the genital apparatus, spines are short, broom brush-shaped, and having 14-16 digits at the tip (Fig. 6). The spines on the postero-ventral body surface are sparsely distributed, and they become spade-shape with smaller numbers of digitations at the tip (Fig. 7).
The dorsal body surface is covered densely with tegumental spines and sensory papillae, except around the posterior end of the body and excretory pore (Fig. 8).
The antero-lateral body surface is covered with protruded cytoplasmic processes, each with a 15-17 pointed tegumental spine (Fig. 9). On the mid-lateral surface, tegumental spines become more densely crowded and wider in shape with 19-21 pointed tips; uni-ciliated sensory papillae are at times observed (Fig. 10). In some areas of the mid-lateral surface, the cytoplasmic processes are protruded, and each process bears a spade-shaped tegumental spine with 12-14 digits (Fig. 11). On the mid-posterior body surface, the tegumental spines are sparse and claw-shaped with 4-5 digits (Fig. 12). On the postero-lateral body surface, spines are also sparsely distributed and claw-shaped with 3-4 digits (Fig. 13). The tegument near the posterior end of the body is severely wrinkled with deep furrows and devoid of spines (Fig. 14).
DISCUSSION
The surface ultrastructures of P. summa were generally similar to that of other heterophyid flukes; M. yokogawai (Lee et al., 1984), M. miyatai (Chai et al., 1998b), M. takahashii (Chai et al., 2000), H. continua (Hong et al., 1991), and H. nocens (Chai et al., 1992). However, significant differences were recognized in the morphology and distribution of tegumental spines and sensory papillae.
In P. summa, well-differentiated spines had 14-16 pointed tips ventrally, and 19-21 pointed tips dorsally. In Metagonimus spp., digitation of the spines was less; 7-9 (ventrally) and 9-11 tips (dorsally) in M. yokogawai (Lee et al., 1984), 9-11 (ventrally) and 10-12 tips (dorsally) in M. miyatai (Chai et al., 1998b), and 10-12 (ventrally) and 10-13 tips (dorsally) in M. takahashii (Chai et al., 2000). In H. continua, tegumental spines on the ventral or dorsal surface are with 15-17 pointed tips (Hong et al., 1991), but less than that in P. summa. In H. nocens, the spines on the ventral surface are 12-17 pointed, and those on the dorsal surface are 15-20 pointed (Chai et al., 1992). Therefore, digitation of the spines of P. summa resembles that of H. nocens.
The shape of tegumental spines is a characteristic feature in the surface ultrastructure of heterophyid flukes. In P. summa, the spines were, in most cases, broom brush- or painting brush-shaped, and in some areas, they were spade-shaped. Near the posterior extremity, the spines were mostly bear-claw-shaped. In Metagonimus spp. and H. nocens, the spines are normally spade- or round brush-shaped (Lee et al., 1984; Chai et al., 1998b, 2000). Broom brush-shaped spines, similar to those observed in this study, have also been observed in H. continua (Hong et al., 1991). However, the presence of painting brush-shaped or bear-claw-shaped spines in P. summa seemed to be a unique feature among different species of heterophyid flukes.
Two types of sensory papillae, type I and type II, were observed in P. summa. These papillae have been found as common features among heterophyid flukes. Type III sensory papillae (round swelling of cytoplasmic ridges), which were observed on the oral sucker of M. yokogawai (Lee et al., 1984), were absent in M. miyatai (Chai et al., 1998b), M. takahashii (Chai et al., 2000) and P. summa (this study).
It is worth to mention here that, light microscopically, minute spines are present on the gonotyl (5 on the right side and 9 on the left side) of the genital apparatus in 7-day adults of P. summa (Chai et al., 1986). We tried to observe the detailed morphology of the spines by scanning electron microscopy, but failed to reveal them, most likely because the gonotyl and the spines were embedded within the genital apparatus.
Changes in the differentiation of tegumental spines in accordance with development of worms have been reported in trematodes. In the case of Clonorchis sinensis, larval flukes which just excysted in the duodenum had double- or triple-pointed tegumental spines at anterior half of the body, but spines gradually disappeared as the worms grew to be adults (Fujino et al., 1979; Lee et al., 1982). On the contrary, in larval flukes of Fasciola hepatica, tegumental spines with a single tip metamorphosed into multipointed ones just prior to entry into the bile duct (Bennett and Threadgold, 1975). Conversion of simple spines into more serrated ones during the parasite development was also found in Neodiplostomum seoulense (Lee et al., 1985) and Paragonimus iloktsuenensis (Lee et al., 1989). Studies on changes in surface ultrastructure of P. summa according to developmental stages are needed.