Introduction
In coastal ecosystems, marine bivalves are frequently associated with various other marine organisms, which may act as either parasites or symbionts [1–3]. Notably, numerous species of single-celled microorganisms and multicellular metazoan parasites have been identified in the Manila clam (Ruditapes philippinarum), a species that dominates the sandy-mud tidal flats along the Yellow Sea and East Sea coasts [4–9]. These parasitic and symbiotic organisms often place considerable stress on their hosts, resulting in both sublethal and lethal effects [10,11]. Among the parasites infecting marine mollusks, the alveolate protist Perkinsus olseni is recognized as a significant pathogen, particularly impacting Manila clams along Korea’s west and south coasts [11–14]. Severe P. olseni infections in Manila clams can lead to reduced reproductive capacity, slower growth rates, and increased mortality [10,12–14]. Therefore, accurate detection and quantification of this pathogen are vital for the effective management and control of P. olseni infections.
Infection by P. olseni has been diagnosed using various assays, including species-specific PCR, histology, and the anaerobic fluid thioglycollate medium (FTM) assay [4,15–17]. To monitor Perkinsus marinus infection in the eastern oyster (Crassostrea virginica), Ray [18] first introduced the use of FTM—a simple and conventional method for parasite detection. During incubation in FTM, P. marinus trophozoites enlarge substantially, increasing from a few microns to as much as 100 μm in diameter, and transform into prezoosporangia (i.e., hypnospores) characterized by thick cell walls. As the walls of encysted prezoosporangia are composed of cellulose, they readily absorb iodine stain, enabling microscopic visualization of prezoosporangia in infected tissues [11,19,20]. To isolate and count these structures from FTM-incubated oyster tissues, Choi et al. [21] developed the NaOH digestion method. In 2M NaOH, the host tissue is fully digested, while P. marinus hypnospores remain intact and can be quantified using a blood cell counter [21]. This RFTM method, combined with 2M NaOH digestion, has since been adapted for the quantitative assessment of P. olseni infection in Manila clams [22–25].
Enumeration of P. olseni cells is carried out using 2 primary approaches: whole-body RFTM assays, which quantify P. olseni from all dissected tissues, and subsample RFTM assays, which estimate infection levels from specific organs, such as the gills [11,12,14]. The gill RFTM assay has been recognized as a practical and reliable proxy for estimating whole-body infection levels. In Isahaya Bay, Japan, P. olseni cell density in clam gill tissue showed a strong positive correlation with total-body cell counts (r2=0.92), whereas infection intensities in other tissues displayed weaker correlations [15]. Similarly, Dang et al. [23] reported a high positive correlation (r2=0.90) between P. olseni cell counts in gill tissue and total-body cell numbers in Manila clams from Arcachon Bay, France.
However, the generalizability of this diagnostic proxy across diverse ecological contexts has not been thoroughly assessed. In particular, low-intensity infections—often marked by limited tissue colonization and reduced parasite detectability—may compromise the diagnostic sensitivity of gill assays. When infections are primarily localized in non-gill tissues, such as the visceral mass (VM), especially during early or mild stages, gill-based detection may produce false negatives. This raises a critical question of whether the gill RFTM assay can reliably detect P. olseni in regions where infection intensity is low.
To address this, the present study examines the tissue-specific distribution of P. olseni in Manila clams collected from 6 geographically distinct tidal flats in Korea. Specifically, we aim to (1) quantify infection intensity across different tissues using the RFTM method, (2) evaluate the correlation between gill-specific and whole-body infection levels under varying infection intensities, and (3) assess the diagnostic reliability of gill tissues as proxies for systemic infection. Our findings aim to clarify the strengths and limitations of gill-based diagnostics in epidemiological surveillance of P. olseni, thereby informing targeted disease monitoring strategies tailored to regional conditions.
Materials and Methods
Sampling sites selection and condition index
Six tidal flats along the west and south coasts of Korea were selected for Manila clam collection in the present study (Supplementary Fig. S1). Site selection was guided by prior data indicating substantial regional variation in P. olseni infection intensity. Specifically, clams from the south coast and from Hwangdo in Cheonsu Bay on the west coast are known to exhibit high infection levels, with intensities exceeding 1 million cells per gram of gill tissue [17,24,25]. In contrast, clams from the Incheon region and Padori in Taean on the west coast typically show much lower infection intensities [17,25]. To assess the diagnostic performance of the gill RFTM assay across a broad spectrum of infection levels, we selected sampling sites that reflect this geographic variation in P. olseni prevalence and intensity (see also Supplementary Table S1 for the administrative districts and coordinates of each site).
For the assay, Manila clams with shell lengths (the longest axis of the shell) over 30 mm were collected. The soft body was removed from the shell and weighed using an electronic balance. The shell was then dried and weighed to the nearest milligram, and the shell length was recorded in millimeters. The condition index (CI) was calculated as the ratio of wet tissue weight to dry shell weight.
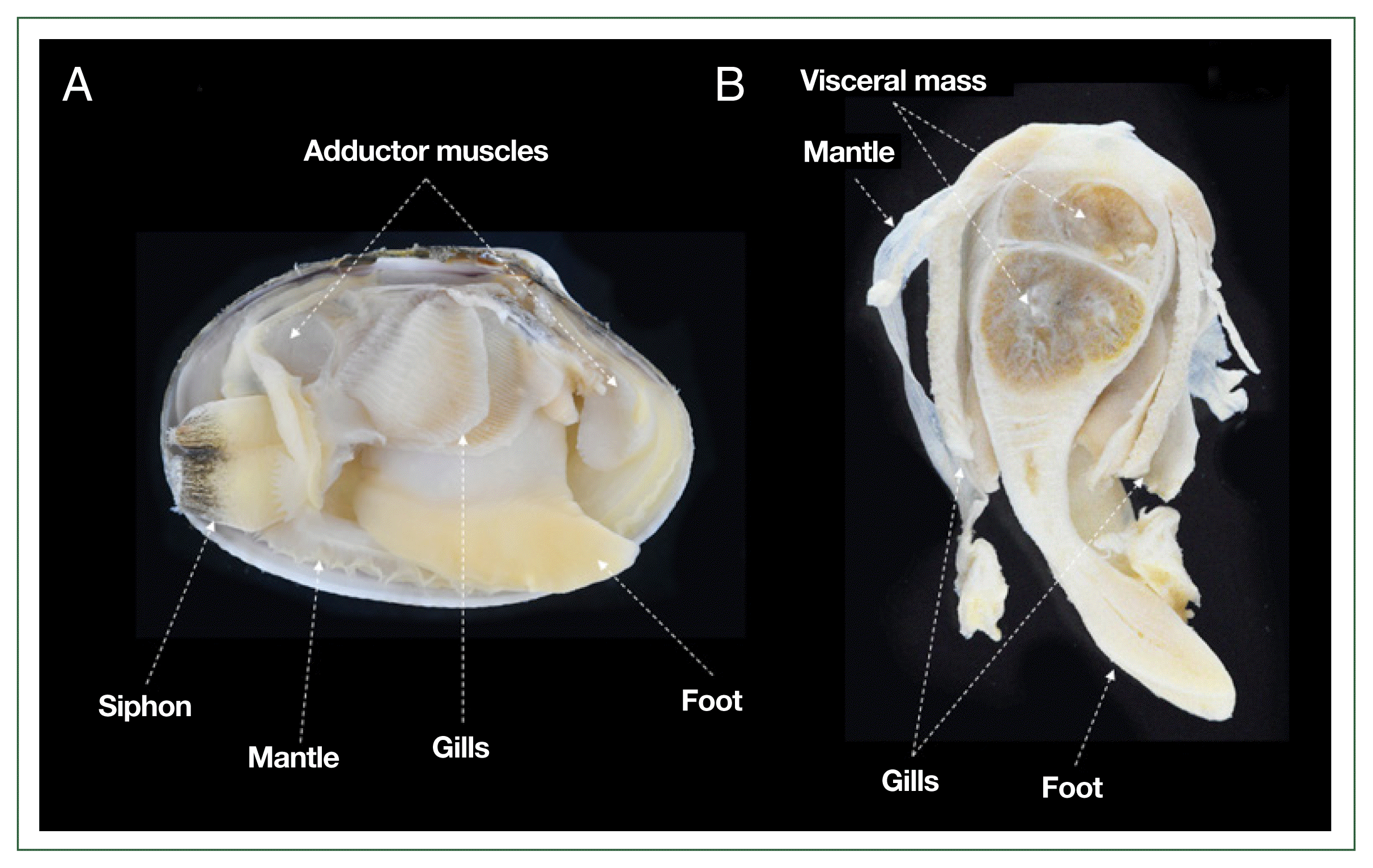
For the RFTM analysis, each clam was aseptically dissected into gills, mantle, adductor muscles, siphon, foot, and VM (Fig. 1). Each tissue was individually weighed to the nearest milligram. Subsequently, the samples were placed in 15 ml tubes containing 10 ml of sterilized FTM, supplemented with nystatin and chloramphenicol to inhibit bacterial growth during incubation [11]. The FTM tubes were incubated in the dark at room temperature for 7 days to induce prezoosporangia formation. After incubation, tissues were sedimented by centrifugation at 2,000×g for 10 min. The supernatant was discarded, and 5 ml of 2 M NaOH was added to the pellet, which was then resuspended. The samples were incubated at 50°C for 1 h to facilitate tissue digestion. The digested tissues were washed several times with 0.15 M phosphate-buffered saline (pH 7.3), and the prezoosporangia were resuspended in a known volume of phosphate-buffered saline. A subsample of this solution was used to count P. olseni prezoosporangia using a hemocytometer, following the method described by Choi et al. [21]. The total number of P. olseni cells per clam was determined by summing counts from all tissues. Infection intensity was expressed as the number of P. olseni cells per gram of tissue.
Statistical analysis
Spatial differences in P. olseni infection intensity among sampling sites were evaluated using a non-parametric Kruskal-Wallis ANOVA, followed by Dunn’s multiple range test for post-hoc comparisons. One-way ANOVA with Tukey’s honestly significant difference test was applied to compare the CI across sites. To assess the relationship between infection intensities in gill tissue and whole-body tissue, linear regression analyses were conducted, and coefficients of determination (r2) were calculated to evaluate the strength of the correlation. Additionally, tissue-specific diagnostic efficacy (%) and false negative rates were calculated to assess the reliability of RFTM assays across different clam tissues. Diagnostic efficacy (%) was defined as the proportion of correctly identified infected individuals among all diagnosed cases, while the false negative rate referred to the percentage of infected individuals incorrectly classified as negative. All statistical analyses were performed using SAS version 9.4 (SAS Institute, Cary, NC, USA), with significance set at P<0.05.
Results
Prevalence and intensity of P. olseni infections
RFTM analysis revealed that most clams examined in this study were infected with P. olseni, as the infection prevalence (i.e., percentage of infected individuals) was 100.0% at most sites, except for the Padori tidal flat, where it was 90.0% (Table 1). The mean total number of P. olseni cells per clam varied considerably across sites. Clams from Hwangdo (4.8×106 cells), Geogumdo (8.8×106 cells), and Masan (2.7×106 cells) harbored more than 1 million P. olseni cells per individual, whereas clams from Jonghyun (0.1×106 cells), Oeri (0.8×106 cells), and Padori (0.3×105 cells) contained fewer than 1 million cells per clam (Table 1).
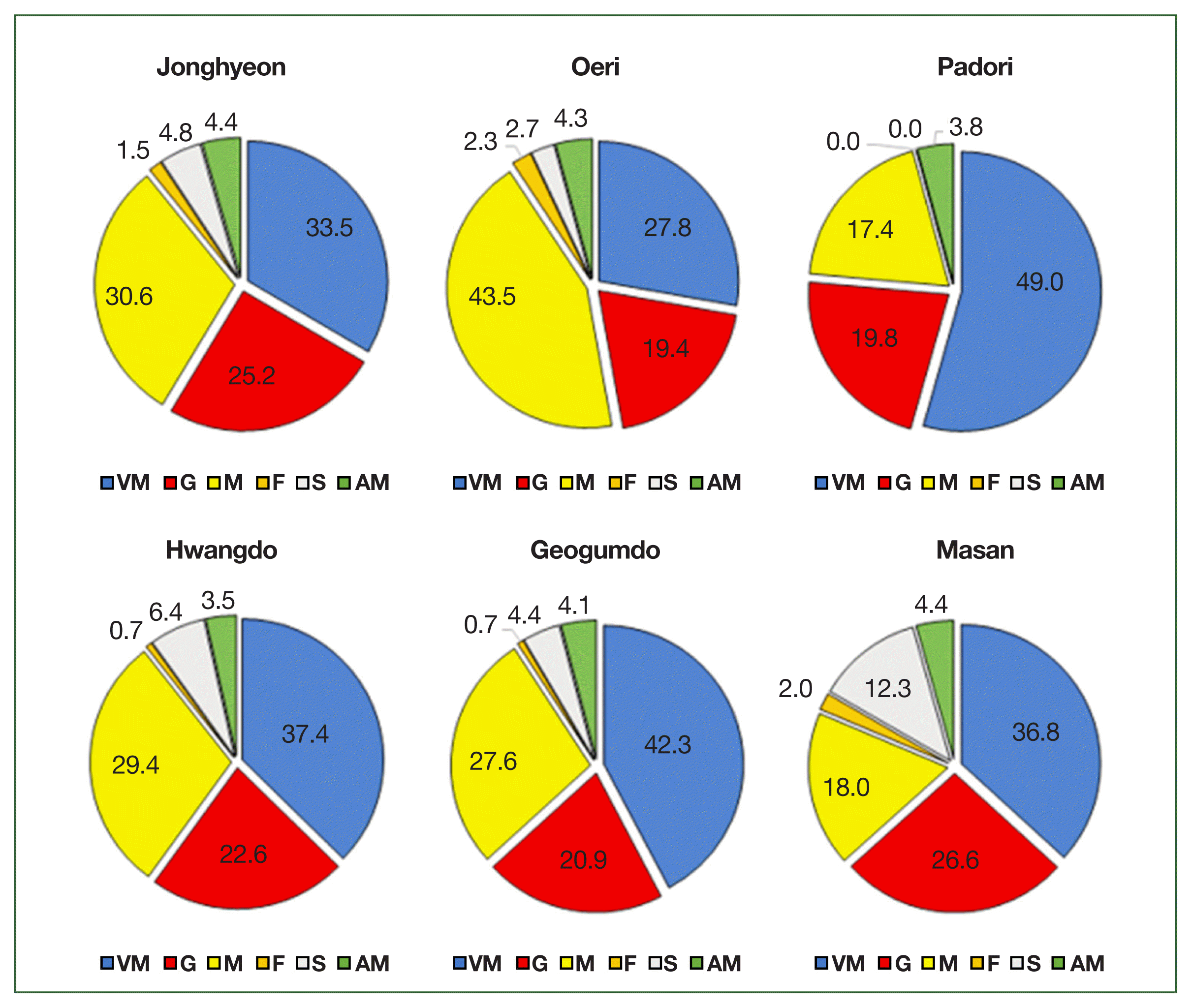
Among the 6 tissue types analyzed, the VM harbored the highest proportion of P. olseni cells, ranging from 27.8% in Oeri to 49.0% in Padori. This was followed by the mantle (M), with proportions ranging from 17.4% in Padori to 43.5% in Oeri, and the gills (G), which ranged from 19.4% in Oeri to 26.6% in Masan (Fig. 2). The foot tissue (F) showed the lowest proportion of P. olseni cells, ranging from 0.0% in Padori to 2.3% in Oeri (Fig. 2).
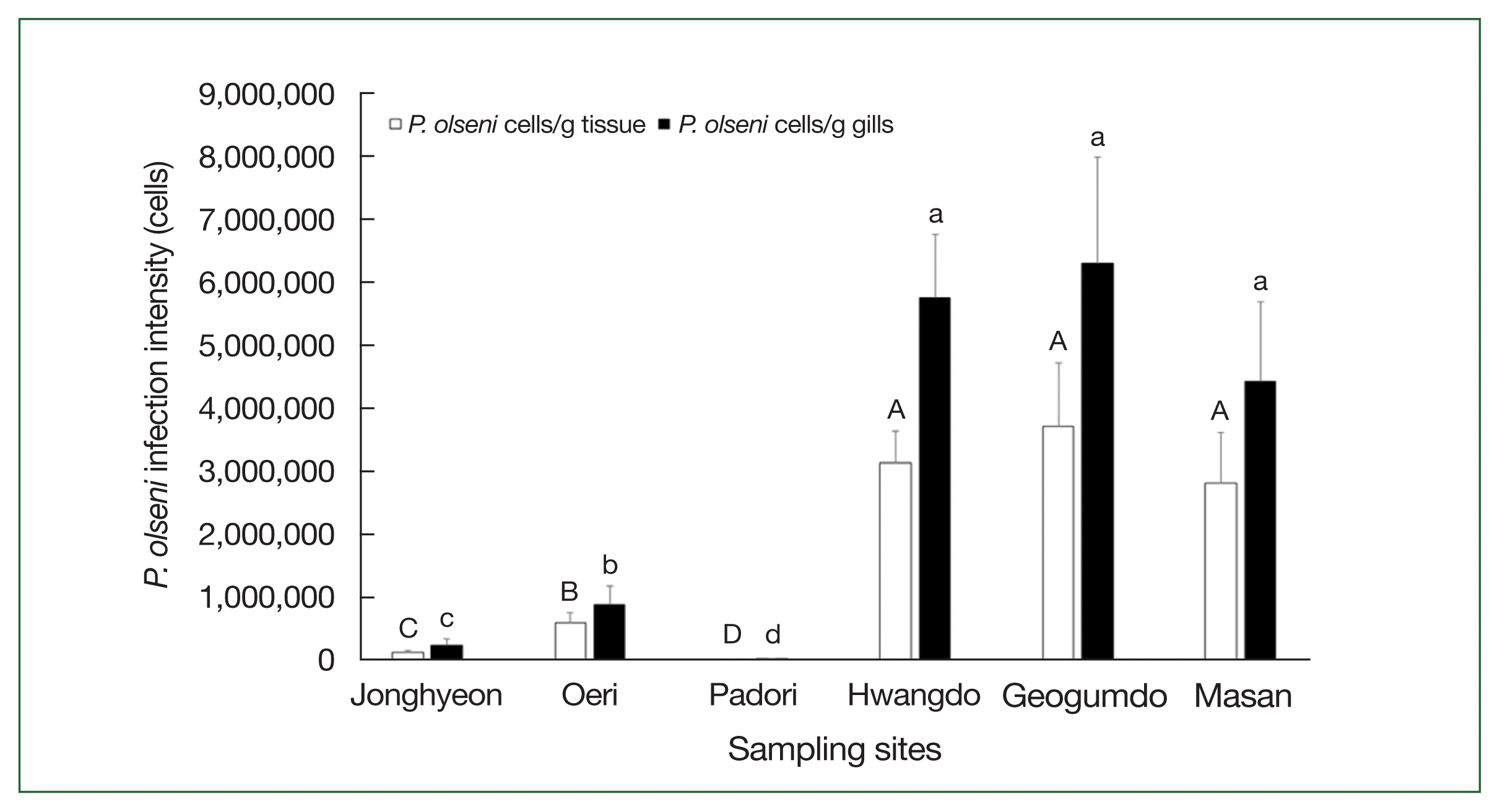
The non-parametric Kruskal-Wallis ANOVA, followed by post-hoc range tests, revealed that P. olseni infection intensities—measured both as cells per gram of total tissue and per gram of gill tissue—were significantly higher (P<0.05) in Hwangdo (3.1×106 cells/g tissue; 5.8×106 cells/g gills), Geogumdo (3.7×106 cells/g tissue; 6.3×106 cells/g gills), and Masan (2.8×106 cells/g tissue; 4.4×106 cells/g gills) compared to those in Jonghyun (0.1×106 cells/g tissue; 0.2×106 cells/g gills), Oeri (0.6×106 cells/g tissue; 0.9×106 cells/g gills), and Padori (0.1×105 cells/g tissue; 0.3×105 cells/g gills) (Fig. 3).
Tissue-specific infection patterns and diagnostic efficacy
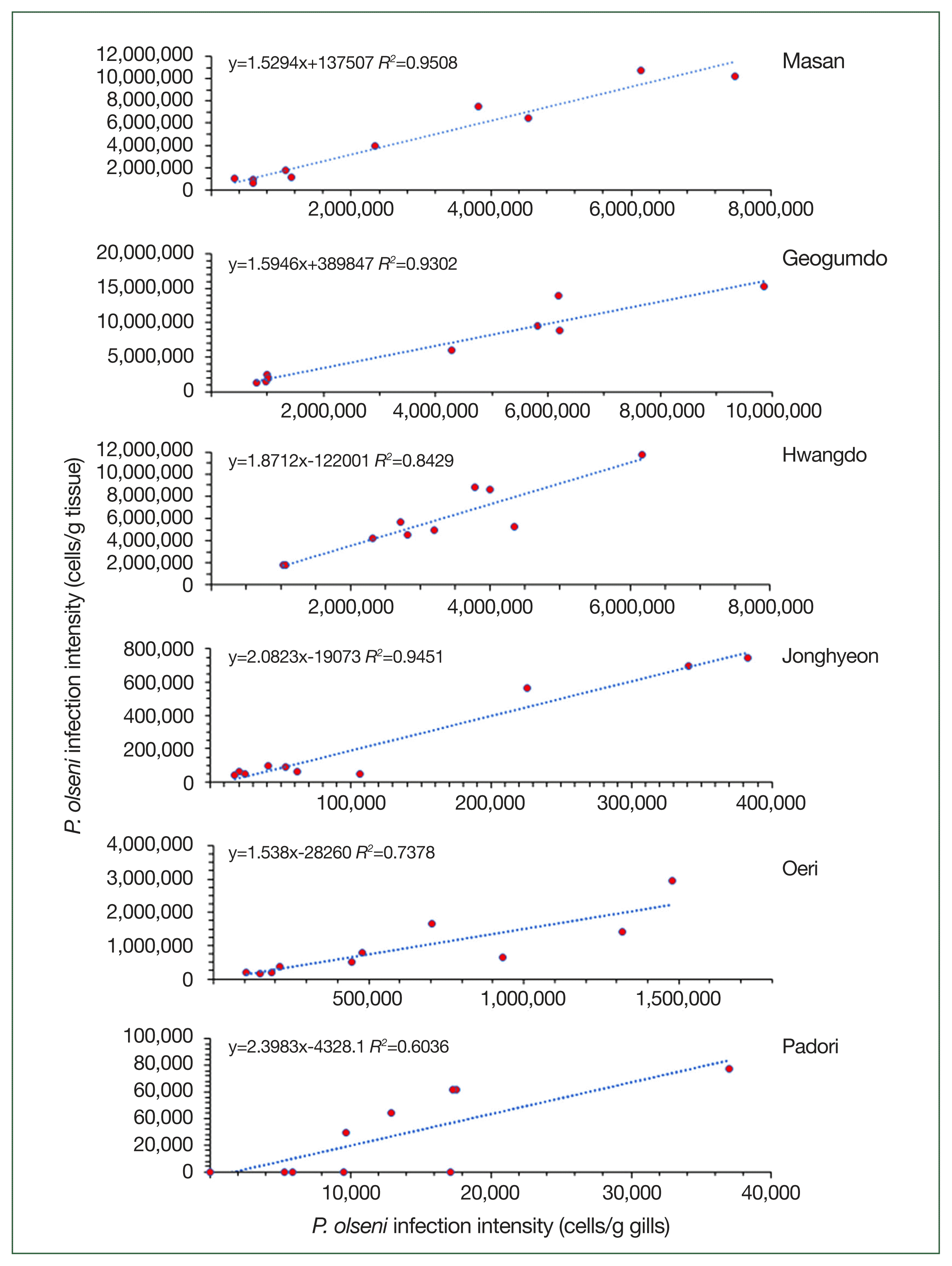
Regression analysis showed a strong positive linear relationship between whole-body RFTM results and gill RFTM results (Fig. 4). Notably, P. olseni cell counts per gram of gill tissue—used as a proxy for estimating individual infection levels—were highly correlated with whole-body infection intensity in Masan (r2=0.951), Geogumdo (r2=0.930), and Hwangdo (r2=0.842), where infection intensities exceeded 1 million cells per gram of tissue. The lowest correlation (r2=0.604) was observed in clams from the Padori tidal flat, which exhibited the lowest infection intensity in this study.
The diagnostic effectiveness of the RFTM assay varied across clam tissues and sampling sites (Table 2). The VM consistently exhibited 100% diagnostic efficacy at all locations, confirming its reliability as the most effective diagnostic tissue. Gill tissue, commonly used as a diagnostic proxy, demonstrated high efficacy at most sites, except for Padori, where the false negative rate reached 80%. Mantle and adductor muscle tissues also showed relatively high efficacy (ranging from 66.7% to 100.0%) but displayed considerable variability across sites—particularly in Padori, where false negative rates were 75.0% and 87.5% for the mantle and adductor muscle, respectively. Siphon and foot tissues showed the lowest diagnostic reliability, with efficacy dropping to 0.0% in Padori.
CI variation among sites
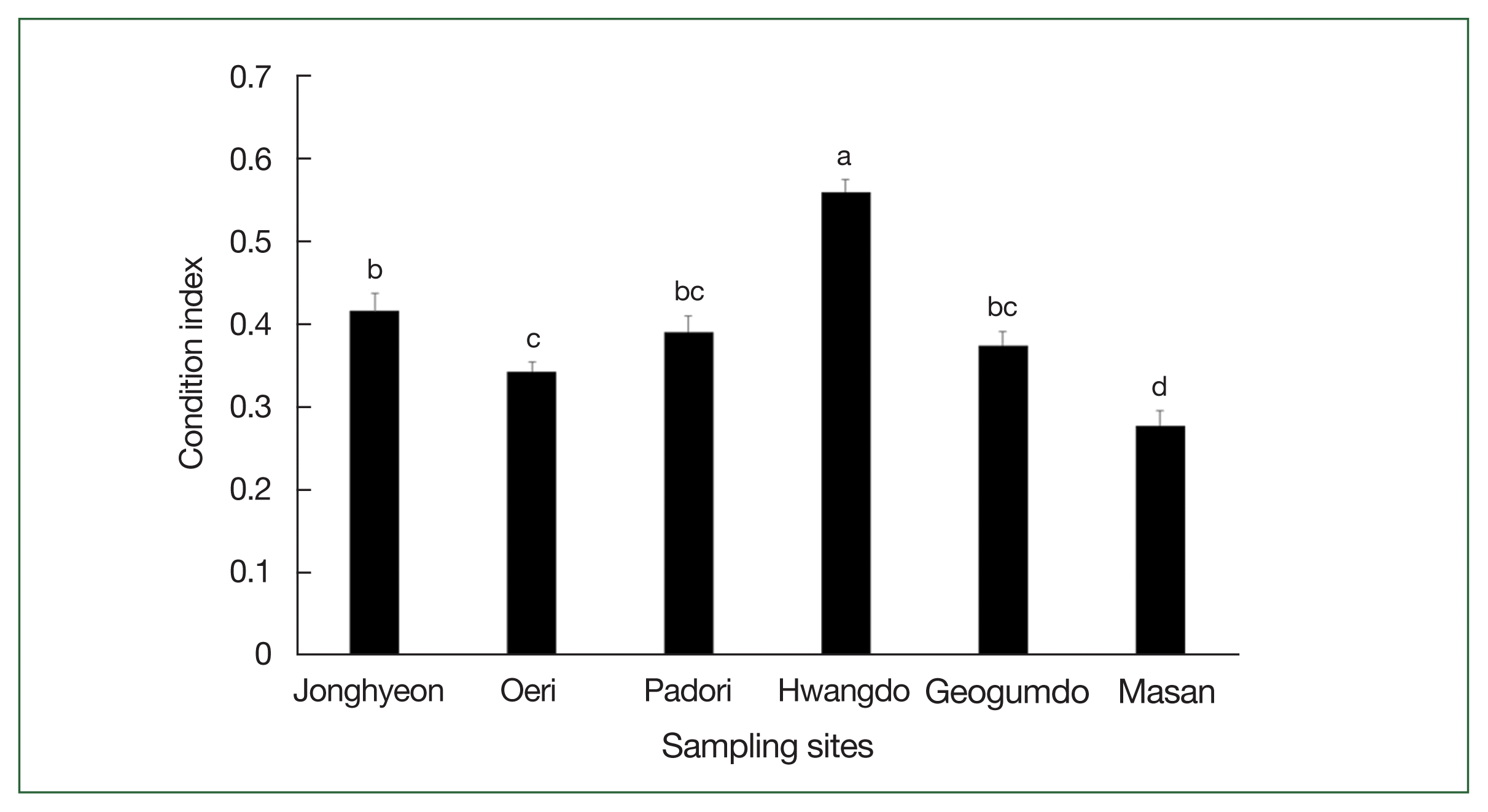
The CI, used as a proxy for clam health, varied substantially among sites, ranging from 0.3 in Masan to 0.6 in Hwangdo (Fig. 5). One-way ANOVA indicated that the mean CI of clams from Hwangdo was significantly higher than those from other tidal flats (P<0.05), while clams from Masan exhibited the lowest mean CI, which was significantly lower than those from the remaining 5 sites (P<0.05).
Discussion
Several diagnostic methods have been employed to detect and quantify P. olseni infections, each offering distinct advantages and limitations. The RFTM assay is a conventional, cost-effective, and widely used method that allows visualization and quantification of enlarged prezoosporangia. However, it requires extended incubation times and lacks species specificity, as it cannot differentiate P. olseni from other Perkinsus species [11,19]. Histological examination enables direct visualization of P. olseni within host tissues, but it is labor-intensive and less sensitive, particularly for detecting low-intensity infections. PCR-based approaches, including species-specific quantitative PCR, offer rapid and highly specific detection and are increasingly used for large-scale surveillance. Nonetheless, PCR methods do not quantify infection burden, are susceptible to sample inhibition, and require more advanced laboratory infrastructure. Considering the trade-offs between practicality and quantitative capacity, the RFTM assay remains a valuable tool, especially for studies aiming to assess infection intensity across tissues and in diverse field conditions.
The tissue-specific distribution of P. olseni revealed that the VM, mantle, and gills consistently harbored the highest parasite loads, while the siphon, foot, and adductor muscles exhibited notably lower infection levels. These findings align with previous studies suggesting that P. olseni preferentially colonizes visceral and digestive tissues [12,23]. In regions such as Padori, where infection intensity was low, the VM demonstrated superior diagnostic efficacy and lower false negative rates compared to gill tissues. This raises an important consideration when selecting tissues for diagnostic purposes.
Although the VM assay showed comparable diagnostic efficacy to whole-body sampling, it is essential to consider the practical implications of tissue selection beyond diagnostic performance. Specifically, the gill assay offers a methodological advantage: by using only a small portion (i.e., One of the 2 pairs of gills is sufficient for the RFTM assay), it preserves the remainder of the specimen for complementary analyses. This enables multi-level evaluations from the same individual, including histopathological assessments (via sections of the VM) and CI measurements. In contrast, the VM comprises a substantial portion of soft tissue, and its use for RFTM analysis limits the ability to conduct parallel histological or physiological assessments. In this study, the gill RFTM assay demonstrated a strong correlation with whole-body infection levels in high-infection areas, supporting its continued use in routine diagnostics. From an integrative research perspective, gill sampling offers greater analytical flexibility without compromising diagnostic accuracy in high-infection contexts.
Nevertheless, our findings also highlight the limitations of relying solely on gill tissue in low-infection environments. In such cases, the sensitivity of the gill assay may decrease, increasing the likelihood of false negatives. To address this, we recommend a flexible, context-specific diagnostic approach: while the gill RFTM assay may be sufficient in high-prevalence areas, a dual-tissue strategy—incorporating both gill and whole-body samples—should be adopted for enhanced diagnostic sensitivity in low-infection zones or during early-stage monitoring. This adaptable framework can improve the accuracy and reliability of P. olseni surveillance across varying ecological conditions.
Our results revealed significant regional variation in P. olseni infection intensity across the 6 tidal flats surveyed. High infection levels were observed in Hwangdo, Geogumdo, and Masan, while relatively low infection intensities were recorded in Padori, Oeri, and Jonghyeon. These geographic differences align with previous studies reporting higher P. olseni burdens in southern coastal regions compared to western regions of Korea [17]. These disparities are likely influenced by ecological and environmental factors, such as temperature, salinity, sediment type, and nutrient availability, which affect both parasite proliferation and host susceptibility [11,12].
The CI of Manila clams varied significantly across sites and appeared to be negatively correlated with P. olseni infections. Clams from Masan, which exhibited high parasite loads, showed the lowest CI values, suggesting a physiological cost associated with infection. Heavy infections can indeed result in reduced reproductive output, impaired growth, increased mortality, and greater susceptibility to secondary infections [10,12]. Chronic infections weaken clams, making them more vulnerable to environmental stressors such as extreme temperatures and hypoxia, which could have broader implications for Manila clam fisheries and aquaculture sustainability. However, it is notable that clams from Hwangdo, despite having high infection intensities, also had the highest CI values, indicating that other factors—such as reproductive stage, food availability, or environmental quality—may also influence clam condition. Therefore, while CI can serve as a useful proxy for host health, it should be interpreted alongside other physiological and biochemical indicators. Ultimately, integrating infection diagnostics with health indices offers a more comprehensive approach to assessing the impact of parasitic infections on the host.
In conclusion, this study offers a thorough evaluation of tissue-specific P. olseni infection patterns and highlights both the strengths and limitations of the gill RFTM assay across different ecological contexts. The results confirm that gill tissue is a reliable proxy for assessing whole-body infection intensity, especially in populations with high infection loads. Moreover, the observed variation in P. olseni prevalence and intensity across sites emphasizes the role of regional environmental factors in shaping infection dynamics. These findings provide valuable insights for developing site-specific surveillance strategies for Manila clam populations, particularly in regions where P. olseni poses a significant threat to both aquaculture and wild populations.