Abstract
Dendritic cells have been known as a member of strong innate immune cells against infectious organelles. In this study, we evaluated the cytokine expression of splenic dendritic cells in chronic mouse toxoplasmosis by tissue cyst-forming Me49 strain and demonstrated the distribution of lymphoid dendritic cells by fluorescence-activated cell sorter (FACS). Pro-inflammatory cytokines, such as IL-1α, IL-1β, IL-6, and IL-10 increased rapidly at week 1 post-infection (PI) and peaked at week 3 PI. Serum IL-10 level followed the similar patterns. FACS analysis showed that the number of CD8α+/CD11c+ splenic dendritic cells increased at week 1 and peaked at week 3 PI. In conclusion, mouse splenic dendritic cells showed early and rapid cytokine changes and may have important protective roles in early phases of murine toxoplasmosis.
-
Key words: Toxoplasma gondii, spleen, dendritic cell, cytokine, FACS, mouse
INTRODUCTION
Toxoplasma gondii is an obligate intracellular protozoan parasite infecting felines as its definitive hosts and an important zoonotic pathogen that can cause severe diseases especially in congenitally infected patients and individuals with human immunodeficiency virus (HIV) patients [
1,
2]. Although
T. gondii is well known to infect macrophages primarily, it is capable of invading and replicating within a wide variety of host cells including dendritic cells.
Dendritic cells (DC) have been known as the major sentinels of the strong innate immune cells against infectious organelles [
3]. Maturation and activation of DC in response to infection plays a major role in the initiation of innate immunity and the subsequent development of adaptive immunity. In toxoplasmosis, it has been known that mice depleted DC are more susceptible to
T. gondii [
4]. This phenomenon has been related with the reduced IL-12 production in the infected mice. Although IFN-γ is the major cytokine of resistance to
T. gondii [
5], other cytokines, such as IL-12, synthesized by DC are also known as crucial triggering cytokines to provoke a series of cell-mediated immunity [
4,
6]. Therefore, in the present study, we investigated on the changes of pro-inflammatory cytokine expression by splenic dendritic cells in chronic murine toxoplasmosis.
MATERIALS AND METHODS
T. gondii ME49 strain
Tissue cyst-forming strain of T. gondii ME49 was passaged with oral route of infection from the previously infected mouse brain of BALB/c strain. Fifty cysts bearing bradyzoites in homogenized mouse brain tissues were introduced orally to experimental groups. A total of 5 mice were used in each group; normal, week 1, 2, 3, 4, and 6 post-infection (PI).
Serum collection and isolation of splenic dendritic cells
After sacrificing mice, serum was collected from each mice and spleen was digested with collagenase/EDTA solution. After a positive magnetic bead reaction using dendritic cell isolation kit (Miltenyi Biotec, Bergisch Gladbach, Germany), the spleen DC were collected by passing over a Midi column (Miltenyi Biotec, Bergisch Gladbach, Germany).
FACS analysis of dendritic cells
Individual splenic DC was digested in collagenase/EDTA solution. To measure the viability of DC, we analyzed the DC population using flow cytometric analysis (FACS Calibur, Becton Dickinson, Mountain View, California, USA) and BD Cell Quest Pro (Ver 5.2) software. The population of DC in the dot plot was gated and confirmed based on the expression of surface DC markers, CD11c molecules. For this, phycoerythrin (PE)-labeled anti-CD11c antibody (anti-mouse, hamster) and fluorescein isothiocyanate (FITC)-labeled anti-CD8α antibody (anti-mouse, rat) were used. Isotype controls of PE-hamster IgG1 and FITC-rat IgG2α were used respectively (all from BD Biosciences).
RT-PCR
RNA was prepared by an RNA easy column (Intron, Seoul, Korea) and primer sets of IL-1α, IL-1β, IL-6, IL-10, and TNF-α were purchased from Bioneer (Cheongwon, Korea) and their primer set is listed in
Table 1. RT-PCR reactions using the 1-step RT premix (Intron, Seoul, Korea) were performed according to the manufacturer's instructions; briefly, with an annealing temperature of 55℃ for 35 cycles. Expression results were calculated relative to the expression in β-actin and were performed in triplicate.
After collection of sera, the IL-10 concentration in the mouse toxoplasmosis was measured using the commercial kit (BioLegend, San Diego, California, USA) according to the manufacturer's instructions and assayed in duplicate.
Statistics
Statistical significance was analyzed by the student's t-test. A P-value of <0.05 was considered significant.
RESULTS
FACS analysis of CD8α+/CD11c+ DC
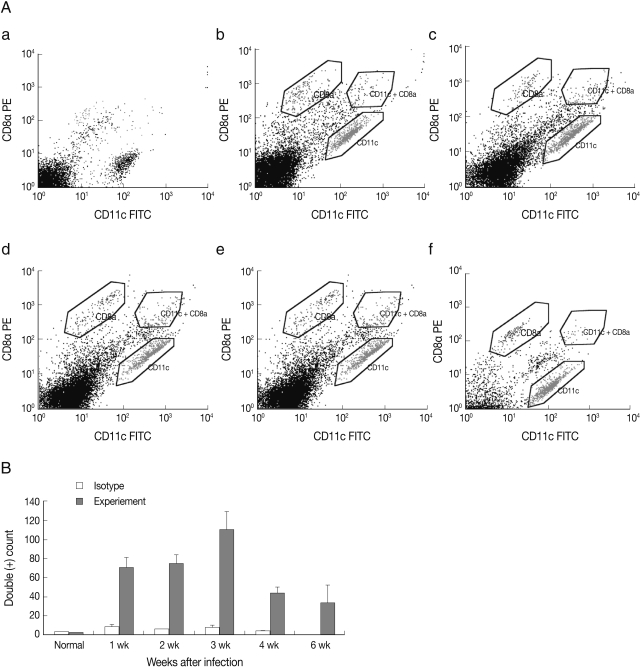
In
Fig. 1, the results of FACS analysis, the number of CD8α
+ CD11c
+ DC was significantly increased at week 1 and peaked at week 3 PI, and thereafter the number of CD8α
+ CD11c
+ DC decreased.
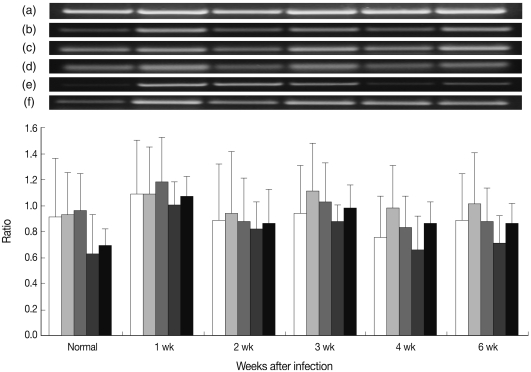
As shown in
Fig. 2, most pro-inflammatory cytokines, such as IL-1α, IL-6, IL-10, and TNF-α increased significantly at week 1 PI. Especially, IL-1β increased at week 1 and peaked at week 3 PI. Expression of all pro-inflammatory cytokines at week 6 PI was maintained higher than in the pre-infected group.
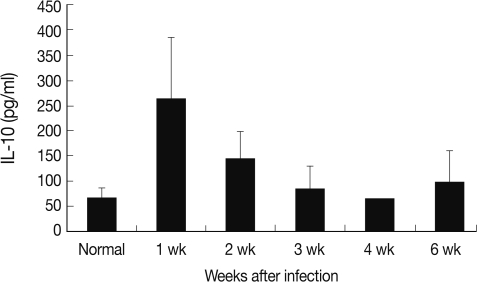
Each cytokine expression of RT-PCR followed similar chronological changes. Therefore, IL-10, which has an important role in DC, was selected and measured its serum levels in mouse toxoplasmosis. The serum level of IL-10 increased significantly at week 1 PI (264.1 pg/ml) than normal (67.4 pg/ml) as in
Fig. 3.
DISCUSSION
DC are well known to be the major sentinels of the innate immune system. Although all DC originate from precursors in the bone marrow, fully matured DC are differentiated into distinct phenotypic subsets. Two major lineages, myeloid and plasmacytoid, are known to develop in mice and humans. Especially in the mouse, myeloid DC are further subdivided into CD8α
+, CD4
+, and CDα
-CD4
- subsets [
7] and lymphoid DC develop from an early lymphoid progenitor under the influence of IL-3 but independent of GM-CSF [
8-
10]. Phenotypic characteristics of these lymphoid DC include their expression of CD8α and CD1d in addition to the pan-DC marker CD11c. These subsets show not only specific immunologic functions but also unique repertoires of pattern recognition receptors, therefore, appears to be capable of responding to various microbial stimuli [
11]. In our experiment, we performed a FACS analysis that CD8α
+ CD11c
+ DC were activated at the early period of week 1 after infection as shown in
Fig. 1A and 1B. These results suggested that CD11c
+ DC in spleen may have important roles both in protective mucosal immunity and propagation of
T. gondii tachyzoites transformed from bradyzoites within cysts after oral infection. It is well known that DC could act as a migrating cell. Therefore, DC have been suggested as a means for some parasites to diffuse into the host [
12,
13]. Also,
T. gondii tachyzoites infected DC can offer as shuttle cells for tachyzoites inducing transmigration into the mesenteric lymph node and spleen [
14,
15].
Recently, CD11c
+ DC in the spleen are known predominantly to be tachyzoites-infected DC after intraperitoneal inoculation of
T. gondii and can transport
T. gondii tachyzoites from the intestinal epithelium to the brain [
14]. Furthermore, these cells are known to show stronger intracellular associations with a certain type II parasite genotype than type I and contribute the tachyzoites dissemination [
16]. In the injection experiments of mice with live tachyzoites or a soluble tachyzoite extract, IL-12p40-producing cells which are consisting entirely of CD11c
+ DC were observed within a few hr in the T-cell areas of the spleen [
6]. In our experiment, staining of the same spleen sections with anti-CD11c monoclonal antibody revealed a massive redistribution of DC in spleen as a consequence of
T. gondii infection, with dominant clustering in the T-cell areas (data not shown). In
Fig. 1 of FACS scan, CD8α
+ CD11c
+ DC is significantly increased at week 1 and peaked at week 3 PI. After then, the number of CD8α
+ CD11c
+ DC decreased. Actually, this decreased period is likely to concord with the term of cyst formation in the brain of infected mice, and in chronic infection, possibly DC from blood monocytes are migrating into the inflammatory sites of perivascular and parenchyma in the central nervous system (CNS) [
17]. Therefore, these results suggested that CD8α
+ CD11c
+ DC might have protective roles in the early phase of murine toxoplasmosis.
In the present study, we compared pro-inflammatory cytokine expression of mouse splenic DC after
T. gondii infection. Pro-inflammatory cytokines, such as IL-1α, IL-1β, IL-6, IL-10, and TNF-α showed a significant increase at week 1 PI as shown in
Fig. 2. Among them, IL-10, known as an anti-inflammatory cytokine, is produced by a variety of cells, including dendritic cells. The IL-10 signaling cascade is a major pathway involved in the control of pro-inflammatory cytokines, such as IL-12 and TNF-α [
18,
19]. Although IL-10 secreted from DC does not appear to be a major mechanism responsible for dampening IL-12 production [
20], the balance between pro-inflammatory cytokine of IL-12 and down regulatory cytokine of IL-10 is known to be essential to control toxoplasmosis [
21]. Actually, IL-10 increased synergistically with the TNF-α expression of splenic dendritic cells at week 1 PI as shown in
Fig. 3.
In conclusion, the present data prove the pro-inflammatory cytokine expression of splenic DC in mice and suggested the possible protective roles of DC in the pathogenesis of toxoplasmosis. Therefore, the roles of some other myeloid related DC in toxoplasmosis should be investigated in the future.
ACKNOWLEDGEMENTS
The authors thank Mr. Je-Young Ryu for his technical assistance. The research was supported by grants from the National Institute of Health (2008-1377-1-1, 2008-E00151-00), Ministry of Health and Welfare, Republic of Korea.
References
- 1. Choi WY, Nam HW, Kwak NH, Huh W, Kim YR, Kang MW, Cho SY, Dubey JP. Foodborne outbreaks of human toxoplasmosis. J Infect Dis 1997;175:1280-1282.
- 2. Dubey JP. Advances in the life cycle of Toxoplasma gondii. Int J Parasitol 1998;28:1019-1024.
- 3. Banchereau J, Briere F, Caux C, Davoust J, Lebecque S, Liu Y, Pulendran B, Palucka K. Immunobiology of dendritic cells. Annu Rev Immunol 2000;18:767-811.
- 4. Liu CH, Fan YT, Dias A, Esper L, Corn RA, Bafica A, Machado FS, Aliberti J. Cutting edge: dendritic cells are essential for in vivo IL-12 production and development of resistance against Toxoplasma gondii infection in mice. J Immunol 2006;177:31-35.
- 5. Suzuki Y, Orellana MA, Schreiber RD, Remington JS. Interferon-gamma: the major mediator of resistance against Toxoplasma gondii. Science 1988;240:516-518.
- 6. Reis e Sousa C, Hieny S, Scharton-Kersten T, Jankovic D, Charest H, Germain RN, Sher A. In vivo microbial stimulation induces rapid CD40 ligand-independent production of interleukin 12 by dendritic cells and their redistribution to T cell areas. J Exp Med 1997;186:1819-1829.
- 7. Shortman K, Liu YJ. Mouse and human dendritic cell subtypes. Nat Rev Immunol 2002;2:151-161.
- 8. Ardavin C, Wu L, Li CL, Shortman K. Thymic dendritic cells and T cells develop simultaneously in the thymus from a common precursor population. Nature 1993;362:761-763.
- 9. Saunders D, Lucas K, Ismaili J, Wu L, Maraskovsky E, Dunn A, Metcalf D, Shortman K. Dendritic cell development in culture from thymic precursor cells in the absence of granulocyte/macrophage colony-stimulating factor. J Exp Med 1996;184:2185-2196.
- 10. Vremec D, Shortman K. Dendritic cell subtypes in mouse lymphoid organs: cross-correlation of surface markers, changes on incubation and differences between thymus, spleen and lymph nodes. J Immunol 1997;159:565-573.
- 11. Manickasingham SP, Edwards AD, Schulz O, Reis e Sousa C. The ability of murine dendritic cell subsets to direct T helper cell differentiation is dependent on microbial signals. Eur J Immunol 2003;33:101-107.
- 12. Moll H, Fuchs H, Blank C, Röllinghoff M. Langerhans cells transport Leishmania major from the infected skin to the draining lymph node for presentation to antigen-specific T cells. Eur J Immunol 1993;23:1595-1601.
- 13. Nargis M, Chisty MM, Ihama Y, Sato H, Inaba T, Kamiya H. Kinetics of Trypanosoma cruzi infection in guinea-pigs, with special reference to the involvement of epidermal Langerhans' cells in the induction of immunity. Parasitology 2001;123:373-380.
- 14. Courret N, Darche S, Sonigo P, Milon G, Buzoni-Gâtel D, Tardieux I. CD11c- and CD11b-expressing mouse leukocytes transport single Toxoplasma gondii tachyzoites to the brain. Blood 2006;107:309-316.
- 15. Lambert H, Hitziger N, Dellacasa I, Svensson M, Barragan A. Induction of dendritic cell migration upon Toxoplasma gondii infection potentiates parasite dissemination. Cell Microbiol 2006;8:1611-1623.
- 16. Lambert H, Vutova PP, Adams WC, Loré K, Barragan A. The Toxoplasma gondii-shuttling function of dendritic cells is linked to the parasite genotype. Infect Immun 2009;77:1679-1688.
- 17. Fischer HG, Bonifas U, Reichmann G. Phenotype and functions of brain dendritic cells emerging during chronic infection of mice with Toxoplasma gondii. J Immunol 2000;164:4826-4834.
- 18. Butcher BA, Kim L, Panopoulos AD, Watowich SS, Murray PJ, Denkers EY. Cutting edge: IL-10-independent STAT3 activation by Toxoplasma gondii mediates suppression of IL-12 and TNF-α in host macrophages. J Immunol 2005;174:3148-3152.
- 19. Deckert-Schlüter M, Buck C, Weiner D, Kaefer N, Rang A, Hof H, Wiestler OD, Schlüter D. Interleukin-10 downregulates the intra-cerebral immune response in chronic Toxoplasma encephalitis. J Neuroimmunol 1997;76:167-176.
- 20. Reis e Sousa C, Yap G, Schulz O, Rogers N, Schito M, Aliberti J, Hieny S, Sher A. Paralysis of dendritic cell IL-12 production by microbial products prevents infection-induced immunopathology. Immunity 1999;11:637-647.
- 21. Aldebert D, Durand F, Mercier C, Brenier-Pinchart M, Cesbron-Delauw MF, Pellowx H. Toxoplasma gondii triggers secretion of interleukin-12 but low level of interleukin-10 from the THP-1 human monocyte cell line. Cytokine 2007;37:206-211.
Fig. 1FACS analysis of CD8α+/CD11c+ spleen DC in mouse toxoplasmosis. (A) (a) normal, (b) week 1, (c) week 2, (d) week 3, (e) week 4, and (f) week 6 PI. (B) Number of CD8α+/CD11c+ splenic DC after T. gondii infection. Note the number of CD8α+/CD11c+ DC increased significantly at week 1 after T. gondii infection.
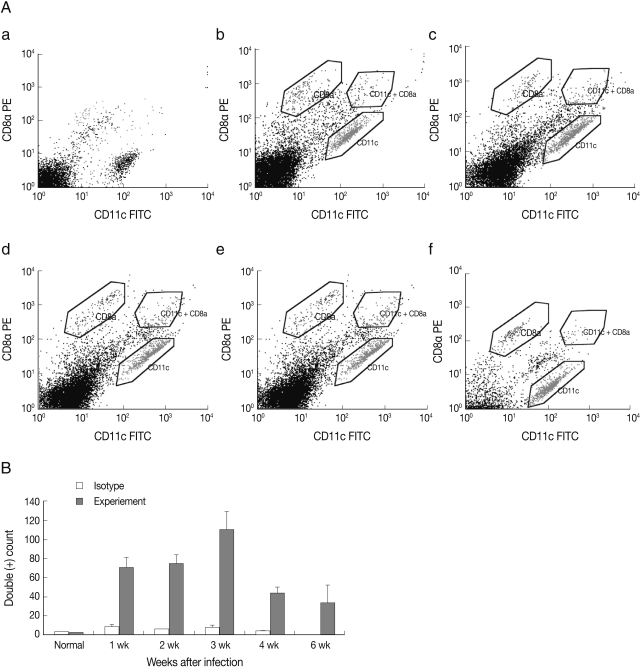
Fig. 2RT-PCR findings of pro-inflammatory cytokines in the splenic DC in mouse toxoplasmosis. Note the increased cytokine expressions at week 1 PI. Each graph shows relative densities against β-actin. (a) β-actin, (b) IL-1α (□), (c) IL-1β (░), (d) IL-6 (▒), (e) IL-10 (▓), and (f) TNF-α (■).

Fig. 3Mouse serum IL-10 levels chronologically in T. gondii infection. The highest IL-10 concentration was measured at week 1 after the infection.
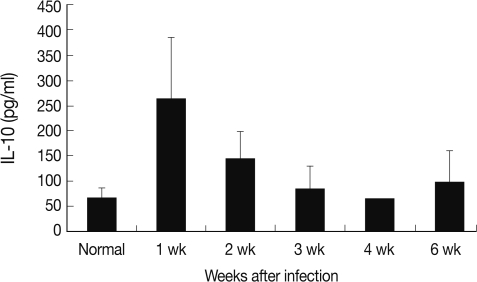
Table 1.Primer sets and corresponding amplicons
Table 1.
|
Name |
Amplicon size (bp) |
|
Primer sequence |
|
β-actin |
395 |
Forward |
5'-AGG CTG TGC TGT CCC TGT ATG C-3' |
|
|
Reverse |
5'-ACC CAA GAA GGA AGG CTG GAA A-3' |
|
IL-1α |
310 |
Forward |
5'-CTC TAG AGC ACC ATG CTA CAG AC-3' |
|
|
Reverse |
5'-TGG AAT CCA GGG GAA ACA CTG-3' |
|
IL-1β |
291 |
Forward |
5'-GCT ACC TGT GTC TTT CCC GTG G-3' |
|
|
Reverse |
5'-TTG TCG TTG CTT GGT TCT CCT TG-3' |
|
IL-6 |
155 |
Forward |
5'-TGG AGT CAC AGA AGG AGT GGC TAA G-3' |
|
|
Reverse |
5'-TCT GAC CAC AGT GAG GAA TGT CCA C-3' |
|
IL-10 |
417 |
Forward |
5'-ATG CAG GAC TTT AAG GGT TAC TTG GGT T-3' |
|
|
Reverse |
5'-ATT TCG GAG AGA GGT ACA AAC GAG GTT T-3' |
|
TNF-α |
300 |
Forward |
5'-GGC AGG TCT ACT TTA GAG TCA TTG C-3' |
|
|
Reverse |
5'-ACA TTC GAG GCT CCA GTG AAT TCG G-3' |