Abstract
Trichomoniasis is a sexually transmitted disease due to infection with Trichomonas vaginalis, and it can cause serious consequences for women's health. To study the virulence factors of this pathogen, T. vaginalis surface proteins were investigated using polyclonal antibodies specific to the membrane fractions of T. vaginalis. The T. vaginalis expression library was constructed by cloning the cDNA derived from mRNA of T. vaginalis into a phage λ Uni-ZAP XR vector, and then used for immunoscreening with the anti-membrane proteins of T. vaginalis antibodies. The immunoreactive proteins identified included adhesion protein AP65-1, α-actinin, kinesin-associated protein, teneurin, and 2 independent hypothetical proteins. Immunofluorescence assays showed that AP65-1, one of the identified immunogenic clones, is prevalent in the whole body of T. vaginalis. This study led us to identify T. vaginalis proteins which may stimulate immune responses by human cells.
-
Key words: Trichomonas vaginalis, antigenic protein, membrane protein, immunoscreening
Trichomonas vaginalis, causing the most prevalent non-viral sexually transmitted disease worldwide, is a popular subject for the study of host-parasite interrelationships [
1]. Infection of the vaginal tract by
T. vaginalis occurs as multiple steps, involving distinct interactions between host macromolecules and protozoan factors. Trichomonads interact specifically with mucin [
2], and then contact vaginal epithelial cells at which iron-regulated surface proteins of
T. vaginalis, adhesion proteins AP65, AP51, AP33, and AP23, play essential roles [
3]. Penetration of the epithelium by
T. vaginalis also induces a specific interaction with extracellular matrix basement membrane glycoproteins [
4].
Iron concentration is a critical factor in the pathogenesis of
T. vaginalis [
5,
6]. The virulence of trichomonads is increased by iron, and iron modulates multiple aspects of
T. vaginalis, including metabolic activity, cytoadherence, and resistance to complement lysis [
5,
6]. Like other pathogens,
T. vaginalis has ways of acquiring iron from lactoferrin [
3]. Upon binding to lactoferrin via specific receptors of
T. vaginalis, this protozoan shows increased iron accumulation and activity of pyruvate ferridoxin oxidoreductase [
7]. Iron availability from lactoferrin triggers trichomonads to increase amount of adhesins, thereby enhancing the binding to the host epithelial cells [
8].
T. vaginalis also binds to erythrocytes which provide both lipid and iron for parasites [
9], and iron acquisition from hemoglobin by
T. vaginalis occurs via the action of 2 adhesins, AP51 and AP65 [
10].
T. vaginalis grown in iron-depleted medium shows no pathology, whereas trichomonads cultured in the iron supplemented medium, could cause subcutaneous abscesses in mice [
11].
In addition to surface proteins, proteins secreted by
T. vaginalis were extensively examined with respect to interaction with human vaginal epithelial cells (VEC) [
12]. The components of
T. vaginalis secreted proteins were identified as metabolic enzymes, proteases, and actin, which induced the expression of host components, including interleukin 8, COX-2, and fibronectin [
12]. In the present study, we investigated on the properties of antigenic proteins present in the membrane portion of
T. vaginalis using immunoscreening to identify virulence factors involved in interaction with host cells.
The Korean
T. vaginalis isolate, KT4 [
11], was used in all experiments. Trichomonads were grown axenically in Diamond's trypticase-yeast extract-iron (TYI-S-33) medium [
13], supplemented with 10% (v/v) heat-inactivated calf serum (Gibco BRL, Rockville, Maryland, USA). Cultures were incubated at 37℃ in a 5% CO
2 atmosphere, and passaged every 2-3 days.
Membrane extraction from
T. vaginalis was carried out as described previously [
14]. Briefly,
T. vaginalis in late logarithmic growth phase was washed 3 times with PBS (137 mM NaCl, 2.7 mM KCl, 10.1 mM Na
2HPO
4, and 2 mM KH
2PO
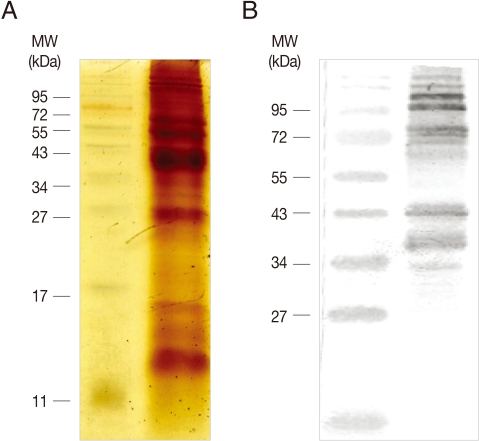
4, pH 7.3), and resuspended in ice-cold PBS containing protease inhibitors (1 mM N-tosyl-L-lysine-chloromethyl ketone hydrochloride, and 0.2 mM leupeptin). Trichomonads were lysed by sonication on ice, and centrifuged for 10 min at 13,000 rpm, 4℃. Membrane proteins were prepared by resuspending the resultant pellet in PBS containing 1% Triton X-100, and stored at -20℃ until further use. Membrane proteins were mixed with 2×SDS-PAGE sample buffer, and boiled for 5 min. After 15% SDS-PAGE electrophoresis, proteins were visualized by staining with silver nitrate (
Fig. 1A).
Membrane proteins prepared from T. vaginalis (100 µg) were mixed with 0.5 ml of complete Freund's adjuvant (Sigma, St. Louis, Missouri, USA), and injected intraperitoneally into a specific pathogen-free, 7-week-old, female rat (CrjBgi:CD[SD]IGS). Two additional immunizations were performed with the same amount of membrane proteins mixed with incomplete Freund's adjuvant (Sigma) at 2 and 4 weeks after the primary immunization. A week after the third immunization, serum was obtained from the immunized rat, and used for western blot analysis.
Membrane proteins of
T. vaginalis (20 µg) were separated by 12% SDS-PAGE, and then transferred to a nitrocellulose filter (Millipore, Billerica, Massachusetts, USA). The membrane was incubated with polyclonal rat anti-membrane protein antibodies in a blocking solution (PBS, 5% skim milk, and 0.05% Tween 20), and then incubated with alkaline phosphatase (AP)-conjugated anti-rat IgG (Sigma). The immunoreactive protein was visualized using the nitroblue tetrazolium (NBT)/5-bromo-4-chloro-3-indolyl phosphate (BCIP) system (Promega, Madison, Wisconsin, USA). As shown in
Fig. 1B, several immunoreactive protein bands were visualized by western blot analysis, indicating that anti-membrane polyclonal antibodies reacted to diverse
T. vaginalis membrane proteins.
To prepare a T. vaginalis cDNA expression library, total RNA was prepared from freshly grown T. vaginalis using TRIzol (Invitrogen, Carlsbad, California, USA), and then processed further to purify mRNA using the PolyATtract mRNA isolation system (Promega). T. vaginalis cDNA was synthesized from this pool of mRNA as a template using a cDNA synthesis kit (Strategene, Santa Clara, California, USA). The resultant cDNAs were ligated with EcoRI and XhoI linkers at their 5'- and 3'-ends, respectively, and then cloned into the corresponding sites of Escherichia coli phage λ Uni-ZAP XR vector (Stratagene). The resultant ligation mixture of T. vaginalis cDNA and Uni-ZAP XR vector was packaged as phage in vitro using Gigapack III gold packaging extract (Stratagene). Twenty plaques were chosen randomly, and their inserts were amplified by PCR, using primers T3 and T7 annealed at both sides of the insert. Agarose gel electrophoresis of the resultant PCR products clearly indicated that most clones of the T. vaginalis library had inserts of various sizes derived from the diverse mRNA of T. vaginalis.
Polyclonal antibodies specific to membrane proteins of
T. vaginalis were used for the immunoscreening experiment. The amplified library (with a titer of 3×10
5 plaque forming units/ml) was screened using a 1:2,000 dilution of rat antibodies raised against
T. vaginalis membrane proteins. Then, 9 plaques, distinct from the background level of antibody binding to the filter, were further purified by second and third screenings. Homogeneous plaques expressing the antigens were selected and excised to a pBluescript SK(+) phagemid, as instructed by the manufacturer (Stratagene). The identities of the phagemid inserts were verified by automatic sequencing of the double-stranded plasmids (
Table 1). The most interesting clone among the isolated cDNAs via immunoscreening was AP65-1, which is 1 of 4 iron-regulated surface proteins involved in the cytoadherence of
T. vaginalis [
15]. The majority of identified clones encode components associated with cytoskeleton, which included actinin, and a kinesin-associated protein. Most notably, 3 of the 9 isolated clones turned out to be α-actinin, the actin-binding and Ca
2+-binding protein with high immunogenicity in
T. vaginalis [
16]. Two of the identified clones encoded a kinesin-associated protein which functions as a regulator for cellular and intracellular movements via interaction with the motor proteins [
17]. One of the clones was also found to encode a putative teneurin, a conserved family of transmembrane proteins involved in intercellular signaling [
18]. Putative amino acid sequences of the remaining 2 clones did not provide any further information after homologous proteins were compared to NCBI databases. AP65 is known as the main adhesin for
T. vaginalis cytoadherence, and is encoded by a multigene family,
ap65-1,
ap65-2, and
ap65-3 [
19]. Therefore, we examined whether AP65-1 is located in the membrane of
T. vaginalis in the subsequent immunofluorescence assay (IFA) using antibodies specific to AP65-1.
A 1,703 bp
ap65-1 DNA fragment amplified with primer AP65-1F-NcoI (5'-CATG
CCATGGGCATGCTTACATCTTCAGTC-3'; underlined bases indicate an NcoI site) and AP65-1R-NotI (5'-GTTA
GCGGCCGCGTAAAGTGGTTCGTAGTC-3'; underlined bases indicate an NotI site) was cloned into pET28b (Novagen, Darmstadt, Germany) to produce plasmid pZhu115. Histidine-tagged recombinant AP65-1 (rAP65-1) was expressed in
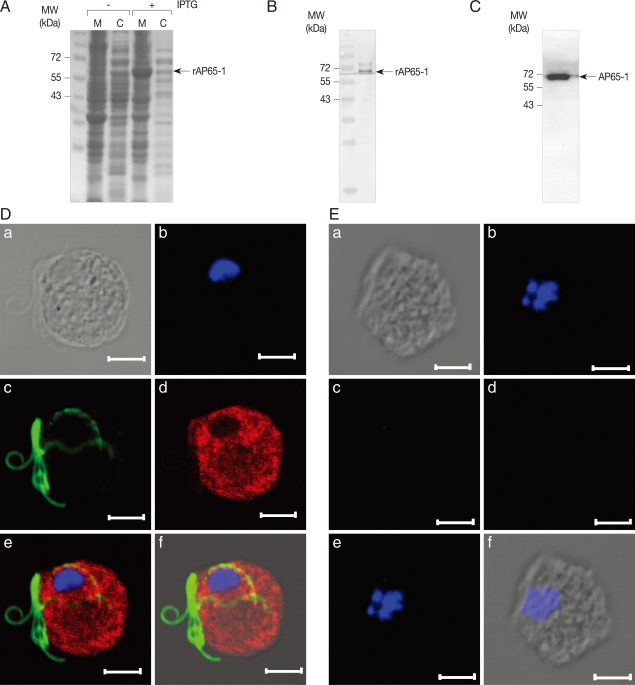
E. coli BL21 (DE3) with an addition of 1 mM isopropyl β-D-thiogalactoside (IPTG) (Sigma), and then used to immunize a specific pathogen-free rat (CrjBgi:CD[SD]IGS, 7-week-old, female) to make anti-rAP65-1 antibodies (
Fig. 2A). Specificity of the resultant antibodies was confirmed by western blot of the
E. coli extracts expressing rAP65-1 (
Fig. 2B). Cell extracts of
T. vaginalis were prepared in PBS, and reacted with AP65-1-specific polyclonal antibodies, clearly indicating an immunoreactive protein of 65 kDa (
Fig. 2C).
The intracellular location of AP65-1 was examined in
T. vaginalis by IFA using the anti-rAP65-1 antibodies (
Fig. 2D). Trophozoites of
T. vaginalis were attached to glass slides coated with L-lysine in a humidified chamber. The attached cells were fixed with chilled 100% methanol at -20℃ for 10 min, and permeabilized with PBS/0.5% Triton X-100 for an additional 10 min. After a 1 hr-incubation in blocking buffer (PBS, 5% goat serum, and 2% BSA), the cells were reacted against rat anti-rAP65 polyclonal antibodies (1:200 dilution) and mouse anti-tubulin antibodies [
20] (1:300 dilution with a blocking buffer) at 4℃ overnight. Following 3 times 5 min-washes with PBS, the cells were incubated with TRITC-conjugated anti-rat IgG and FITC-conjugated anti-mouse IgG (1:200 dilution in blocking buffer, Jackson ImmunoResearch Lab, West Grove, Pennsylvania, USA) at 37℃ for 1 hr. The slides were reacted with 1 µg/ml 4'6-diamidino-2-phenylindole, DAPI (Sigma), rinsed with PBS, and mounted with anti-fade mounting medium (Vectashilde; Vector, Burkingame, California, USA). They were then observed with a Zeiss LSM 510 laser scanning confocal microscope (Zeiss, Thornwood, New York, USA). The images were collected from serial sections at 1 µm intervals, and analyzed by Zeiss LSM image browser software (Zeiss). IFA of
T. vaginalis using anti-rAP65-1 antibodies showed strong red florescence present in the whole body of
T. vaginalis, whereas flagella were stained with green fluorescence, indicating that the intracellular level of AP65-1 is high in cytoplasm and possibly in the membrane of
T. vaginalis.
Representative adhesins of
T. vaginalis, AP65, AP51, and AP33, are found to share sequence homology with metabolic enzymes, which include malic enzymes [
21], succinyl-CoA synthetase β subunit [
22], and succinyl-CoA synthetase α subunit [
23], respectively. It is not unusual that some metabolic enzymes of pathogens play an additional function in interaction with the host; for example, streptococcal surface dehydrogenase working as an adhesin for pharyngeal cells [
24]. Isolation of AP65 as a membrane protein is contradictory with the result of Garcia and Alderete [
15], in which AP65 was found to be secreted and played an important role in
T. vaginalis binding to the host VECs. Therefore, they suggested a model in which AP65, a secreted protein, mediated
T. vaginalis-VEC binding by bridging the 2 receptors, each of which are derived from the parasite and host cells. To resolve this difference, fine fractionation of proteins of
T. vaginalis should be performed in the future experiment.
In conclusion, antigenic membrane proteins were identified by immunoscreening of the T. vaginalis cDNA expression library with anti-T. vaginalis membrane protein antibodies. Future experiments will be performed to define their roles in T. vaginalis-host interaction.
ACKNOWLEDGEMENTS
This work was supported by the Korean Research Foundation Grant funded by the Korean Government (MOEHRD, Basic Research Promotion Fund) (KRF-2007-314-E00070). This work was partially supported by the Seoul R&BD Program (grant 10580), Korea.
References
- 1. Van der Pol B. Trichomonas vaginalis infection: the most prevalent nonviral sexually transmitted infection receives the least public health attention. Clin Infect Dis 2007;44:23-25.
- 2. Lehker MW, Sweeney D. Trichomonad invasion of the mucous layer requires adhesins, mucinases, and motility. Sex Transm Infect 1999;75:231-238.
- 3. Lehker MW, Alderete JF. Iron regulates growth of Trichomonas vaginalis and the expression of immunogenic trichomonad proteins. Mol Microbiol 1992;6:123-132.
- 4. Crouch ML, Alderete JF. Trichomonas vaginalis interactions with fibronectin and laminin. Microbiology 1999;145:2835-2843.
- 5. Tsai CD, Liu HW, Tai JH. Characterization of an iron-responsive promoter in the protozoan pathogen Trichomonas vaginalis. J Biol Chem 2002;277:5153-5162.
- 6. Garcia AF, Chang TH, Benchimol M, Klumpp DJ, Lehker MW, Alderete JF. Iron and contact with host cells induces expression of adhesins on surface of Trichomonas vaginalis. Mol Microbiol 2003;47:1207-1224.
- 7. Peterson KM, Alderete JF. Iron uptake and increased intracellular enzyme activity follow host lactoferrin binding by Trichomonas vaginalis receptors. J Exp Med 1984;160:398-410.
- 8. Lehker MW, Arroyo R, Alderete JF. The regulation by iron of the synthesis of adhesins and cytoadherence levels in the protozoan Trichomonas vaginalis. J Exp Med 1991;174:311-318.
- 9. Lehker MW, Chang TH, Dailey DC, Alderete JF. Specific erythrocyte binding is an additional nutrient acquisition system for Trichomonas vaginalis. J Exp Med 1990;171:2165-2170.
- 10. Ardalan S, Lee BC, Garber GE. Trichomonas vaginalis: the adhesin AP51 and AP65 bind heme and hemoglobin. Exp Parasitol 2009;121:300-306.
- 11. Ryu JS, Choi HK, Min DY, Ha SE, Ahn MH. Effect of iron on the virulence of Trichomonas vaginalis. J Parasitol 2001;87:457-460.
- 12. Kucknoor AS, Mundodi V, Alderete JF. The proteins secreted by Trichomonas vaginalis and vaginal epithelial cell response to secreted and episomally expressed AP65. Cell Microbiol 2007;9:2586-2597.
- 13. Diamond LS. The estabilishment of various trichomonads of animals and man in axenic cultures. J Parasitol 1957;43:488-490.
- 14. Moreno-Brito V, Yañez-Gómez C, Meza-Cervantez P, Avila-González L, Rodríguez MA, Ortega-López J, González-Robles A, Arroyo R. A Trichomonas vaginalis 120 kDa protein with identity to hydrogenosome pyruvate: ferridosin oxidoreductase is a surface adhesion induced by iron. Cell Microbiol 2005;7:245-258.
- 15. Garcia AF, Alderete JF. Characterization of the Trichomonas vaginalis surface-associated AP65 and binding domain interacting with trichomonads and host cells. BMC Microbiol 2007;7:116.
- 16. Addis MF, Rappelli P, Delogu G, Carta F, Cappiccinelli P, Fiori PL. Cloning and molecular characterization of cDNA clone coding for Trichomonas vaginalis alpha-actinin and intracellular localization of the protein. Infect Immun 1998;66:4924-4931.
- 17. Manning BD, Snyder M. Drivers and passengers wanted! The role of kinesin-associated proteins. Trends Cell Biol 2000;10:281-289.
- 18. Tucker RP, Chiquet-Ehrismann R. Teneurins: a conserved family of transmembrane proteins involved in intercellular signaling during development. Dev Biol 2006;290:237-245.
- 19. O'Brien JL, Lauriano CM, Alderete JF. Molecular characterization of a third malic enzyme-like AP 65 adhesin gene of Trichomonas vaginalis. Microb Pathog 1996;20:335-349.
- 20. Kim J, Shin MH, Song KJ, Park SJ. Evaluation of α-tubulin as an antigenic and molecular probe to detect Giardia lamblia. Korean J Parasitol 2009;47:287-291.
- 21. Alderete JF, O'Brien JL, Arroyo R, Engbring JA, Musatovova O, Lopez O, Lauriano CM, Nguyen J. Cloning and molecular characterization of two genes encoding adhesion proteins involved in Trichomonas vaginalis cytoadherence. Mol Microbiol 1995;17:69-83.
- 22. Alderete JF, Engbring J, Lauriano CM, O'Brien JL. Only two of the Trichomonas vaginalis triplet AP51 adhesins are regulated by iron. Microb Pathog 1998;24:1-16.
- 23. Engbring JA, Alderete JF. Three genes encode distinct AP33 proteins involved in Trichomonas vaginalis cytoadherence. Mol Microbiol 1998;28:305-313.
- 24. Pancholi V, Fischetti VA. Cell-to-cell signaling between group A streptococci and paryngeal cells. Role of streoptococcal surface dehydrogenase (SDH). Adv Exp Med Biol 1997;418:499-504.
Fig. 1Preparation of T. vaginalis membrane proteins and formation of anti-T. vaginalis antibodies. (A) T. vaginalis membrane proteins were prepared in PBS/1% Triton X-100, and separated by 15% SDS-PAGE. (B) 20 µg of the membrane proteins were used to immunize rats. Reactivity of the resultant immune sera was examined by immunoblot analysis of T. vaginalis membrane proteins separated by 12% SDS-PAGE.
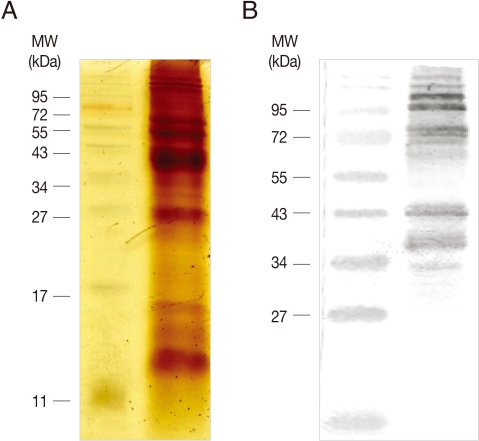
Fig. 2Immunolocalization of AP65-1 adhesin in T. vaginalis trophozoites. (A) Expression of rAP65-1 in E. coli BL21 (DE3). E. coli expressing rAP65-1 (with 1 mM IPTG), was lysed by sonication, fractionated into cytoplasmic (C) and membrane (M) proteins. The resultant proteins were separated by 12% SDS-PAGE. (B) Reactivity of anti-AP65-1 antibodies against E. coli extracts expressing rAP65-1. (C) Detection of AP65-1 in T. vaginalis extracts by western blot analysis using anti-rAP65-1 antibodies. (D) T. vaginalis incubated with rat anti-rAP65-1 antibodies and mouse anti-tubulin antibodies. The slides were then incubated with TRITC-conjugated anti-rat IgG and FITC-conjugated anti-mouse IgG as secondary antibodies. To visualize nuclei, the cells were treated with 1 µg/ml-1 4'6-diamidino-2-phenylindole, mounted with an anti-fade mounting medium, and then observed with an immunofluorescence microscope. The bars represent 5 µm. (E) T. vaginalis trophozoites incubated with rat pre-immune serum. (a) a differential interference contrast (DIC) image, (b) a fluorescence image at a wavelength of 345 nm to detect nuclei, (c) a fluorescence image at a wavelength of 494 nm to detect flagella, (d) a fluorescence image at a wavelength of 547 nm to detect rAP65-1, (e) a combined fluorescence image, and (f) a combined fluorescence and DIC image.
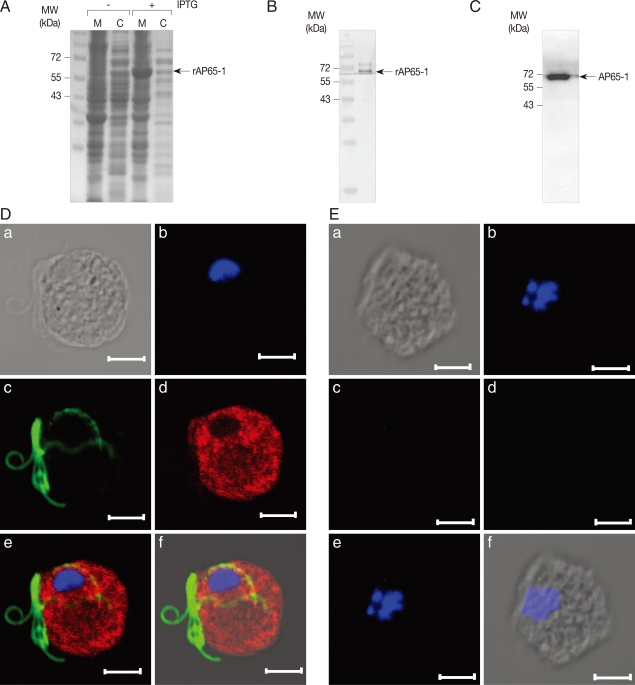
Table 1.Proteins identified by immunoscreening of T. vaginalis cDNA library using anti-T. vaginalis antibodies
Table 1.
|
Clone no. |
T. vaginalis database ORF no. |
NCBI accession no. |
Putative protein |
|
1, 7 |
TVAG_311270 |
XM_001303428 |
kinesin-associated protein |
|
2 |
TVAG_340290 |
U18346 |
AP65-1 adhesin |
|
3, 8, 9 |
TVAG_190450 |
AF072678 |
α-actinin |
|
4 |
TVAG_335250 |
XM_001297646 |
hypothetical protein |
|
5 |
TVAG_402650 |
XM_001580945 |
hypothetical protein |
|
6 |
TVAG_457850 |
XM_001312023 |
teneurin |