Abstract
Clonorchis sinensis is one of the most prevalent parasitic helminths in Korea. Although cholangiocarcinoma can be induced by C. sinensis infection, the underlying mechanism is not clearly understood. To assess the role of C. sinensis infection in carcinogenesis, an in vitro system was established using the human epithelial cell line HEK293T. In cells exposed to the excretory/secretory products (ESP) of C. sinensis and the carcinogen dimethylnitrosamine (DMN), cellular proliferation and the proportion of cells in the G2/M phase increased. Moreover, the expression of the cell cycle proteins E2F1, p-pRb, and cyclin B was dramatically increased when ESP and DMN were added together. Similarly, the transcription factor E2F1 showed its highest level of activity when ESP and DMN were added simultaneously. These findings indicate that DMN and ESP synergistically affect the regulation of cell cycle-related proteins. Our results suggest that exposure to C. sinensis and a small amount of a carcinogen such as DMN can promote carcinogenesis in the bile duct epithelium via uncontrolled cellular proliferation and the upregulation of cell cycle-related proteins.
-
Key words: Clonorchis sinensis, cell cycle, excretory/secretory products, carcinogenesis, dimethylnitrosamine
INTRODUCTION
Clonorchis sinensis is a prevalent parasitic helminth in humans that is endemic in East Asia, including Korea. The main histological features of
C. sinensis infection are irregular dilatation of the bile ducts and glandular hyperplasia [
1-
4]. Over time, infection with
C. sinensis can lead to cholangiocarcinoma; therefore,
C. sinensis is classified as a carcinogen [
3]. Hamsters that are infected with liver flukes such as
C. sinensis or
Opisthorchis viverrini are at the greater risk of developing cholangiocarcinoma through dimethylnitrosamine (DMN)-induced or inflammation-mediated carcinogenesis [
5,
6]. When the bile duct epithelial cells of an infected animal are exposed to high concentrations of
N-nitroso compounds, neoplastic transformation may result [
5-
7]. The
N-nitroso compounds cause nitrosative and oxidative damage to nucleic acids, which may participate in the initiation and/or promotion of cholangiocarcinogenesis [
8]. Using this phenomenon, DMN is commonly used as an initiator of carcinogenesis in rodent models. DNA damage and reparative cellular replication examined to confirm the effects of chemicals such as DMN on hepatocytes [
8,
9].
The mechanisms underlying the proliferation of somatic cells are regulated by cell cycle-related proteins, including cyclins, CDKs, CDK inhibitors, retinoblastoma (Rb) family proteins, and E2F transcription factors [
10]. Cell division is a tightly regulated process that is influenced by innate cues and environmental signals that ultimately control gene expression, post-translational modification, and the proteolysis of multiple regulatory proteins [
11,
12].
Rb proteins function as cancer suppressors by controlling cellular growth through their interaction with E2F and the cyclin A-CDK2 kinase complex [
12,
13]. In fact, the Rb-E2F pathway is one of the most important regulatory pathways controlling cellular proliferation and differentiation in animals and plants [
14,
15]. A number of studies have demonstrated the role of E2Fs in controlling the expression of genes that regulate the G2/M phase of the cell cycle, as well as cell cycle progression and DNA replication [
16]. In particular, the control of
cdc2 and
cyclin B1 transcription is mediated by the interaction of E2Fs with endogenous and G2/M-regulated promoters [
16-
19]. E2F1 is also a key factor in the development of cancer because a number of proto-oncogenes are regulated by E2Fs [
20,
21].
Although the induction of cholangiocarcinoma by
C. sinensis infection and DMN treatment has been demonstrated in a hamster model [
6], the cascade involved in carcinogenesis is unknown. We examined the proliferative effects of excretory/secretory products (ESP) from
C. sinensis and of DMN on human epithelial cells
in vitro.
In cells treated with ESP and DMN, cellular proliferation and the proportion of cells in G2/M phase were increased. Further, the activity of the transcription factor E2F1 was highest when the ESP and DMN were added simultaneously. Our results indicate that ESP and DMN synergistically affect the regulation of cell cycle-related proteins.
MATERIALS AND METHODS
Preparation of C. sinensis excretory/secretory products
To prepare the ESP, metacercariae of C. sinensis were collected from naturally infected freshwater fish (Pseudorasbora parva) at Gyungsangnam-do, an endemic site in Korea. Pepsin-HCl was used to digest the flesh of the fish to obtain the metacercariae, which were then introduced into 6-wk-old Sprague-Dawley rats. After 2 months, C. sinensis adults were collected from the bile ducts of the rats and washed several times with phosphate-buffered saline (PBS) containing 100 µg/ml penicillin and 100 U/ml streptomycin. The freshly isolated worms were then incubated in sterile PBS containing antibiotics for 24 hr in an atmosphere of 5% CO2 at 37℃. After incubation, the medium was centrifuged for 10 min at 800 rpm to remove the worms and debris. The supernatant was then further centrifuged for 10 min at 3,000 rpm and filtered with a syringe-driven 0.45-µm filter unit. The amount of protein in each extract was measured using the Bradford assay (Bio-Rad, Hercules, California, USA).
Cell culture and experimental groups
Human embryonic kidney cells (HEK293T) were cultured in Dulbecco's modified Eagle's medium (DMEM) (Gibco, Rockville, Maryland, USA) supplemented with 10% heat-inactivated fetal bovine serum (FBS) (Gibco), 2 mM L-glutamine, 100 µg/ml penicillin, and 100 U/ml streptomycin at 37℃ in a humidified atmosphere of 5% CO2. Based on the hypotheses to be tested, the cells were divided into 4 groups and cultured for 72 hr: control, cultured in plain medium; DMN, cultured in medium containing 10 µg/ml DMN; ESP, cultured in medium containing 10 µg/ml ESP; and DMN + ESP, cultured in medium containing 10 µg/ml each DMN and ESP.
Antibodies
Polyclonal or monoclonal antibodies were used to detect the expression of several cell-cycle-related proteins. All antibodies were purchased from Santa Cruz Biotechnology (Santa Cruz, California, USA) and used at 1 : 1,000 dilution, including anti-cyclin E (sc-247), anti-cyclin B1 (sc-245), anti-E2F1 (sc-193), and anti-CDK2 (sc-163). Antibodies against calnexin (BD 610523) were purchased from Transduction Laboratories (BD Biosciences, Mountain View, California, USA) and used at 1 : 2,000 dilution. Anti-mouse, anti-rabbit, and anti-goat IgG antisera conjugated with horseradish peroxidase (HRP) were purchased from DAKO (Glostrup, Denmark).
Cellular proliferation assay
We used the XTT formazan method to evaluate the degree of cellular proliferation. For each assay, cells were seeded at a density of 5 × 10
3 cells/well on 96-well plates. After 24 hr of incubation, the medium was replaced with 2% FBS-RPMI 1640 without phenol red. The cells were then incubated in the presence of PBS (vehicle) or 10 µg/ml ESP with or without 10 µg/ml DMN for another 72 hr. XTT (1 mg/ml) was dissolved in warm medium (without phenol red), and 1.25 mM phenazine methosulfate (PMS) was prepared in PBS. Following the incubation of the cells for the indicated periods, 50 µl of the XTT-PMS mixture (final concentration of XTT = 0.3 mg/ml) was added to each well. The plates were then incubated for 4 hr. The conversion of XTT to formazan was quantified by measuring the absorbance at 492 and 690 nm using a microtiter plate reader [
17].
For Western blots, cells were lysed using 1% Nonidet P-40 in a buffer containing 150 mM NaCl, 10 mM NaF, 1 mM PMSF, 200 µM Na3VO4, and 50 mM HEPES, pH 7.4. Equal amounts of protein were separated by 8 and 10% SDS-PAGE and transferred to polyvinylidene fluoride (PVDF) membranes (Immobilon) (Millipore, Bedford, Massachusetts, USA). The membranes were then probed with antibodies against E2F1, CDK2, cyclin B1, pRb, p-pRb, cyclin E, and calnexin. The primary antibodies were detected using goat anti-rabbit or rabbit anti-mouse secondary antibodies conjugated with HRP and visualized using an enhanced chemiluminescence kit (ECL) (Amersham Pharmacia Biotech, Buckinghamshire, UK).
Cell cycle analysis
For cell cycle analysis, HEK293T cells were plated in 6-well culture plates at 2 × 105 cells/well in 2 ml of DMEM containing 10% FBS. They were then treated with 10 µg/ml ESP with or without 10 µg/ml DMN for 72 hr and stained with propidium iodide (PI). The PI-stained cells were analyzed using a FACSCalibur multicolor flow cytometer (BD Biosciences), and the data were analyzed using CellQuest software (BD Biosciences).
Luciferase assay
To analyze the transcriptional activity of E2F1, HEK293T cells (1 × 10
6 cells per 60-mm plate) were transfected with either 10 µg of pXP2-luciferase as a reporter gene (control) or a dihydrofolate reductase (
dhfr) promoter-driven luciferase reporter construct [
22]. Transfection was performed using Lipofectamine™ 2000 (Invitrogen, Carlsbad, California, USA), and the transfected cells were treated with 10 µg/ml ESP with or without 10 µg/ml DMN for an additional 72 hr. Cell extracts were then prepared using 1 × passive lysis buffer (Promega, Madison, Wisconsin, USA), and luciferase activity was measured over 30 sec using a luciferase assay system and luminometer (Turner Designs TD-20/20) (Promega).
Data were expressed as mean value ± standard deviation. The differences between control and the other groups were assessed by the nonparametric Mann-Whitney U test. P < 0.05 was considered to be significant.
RESULTS
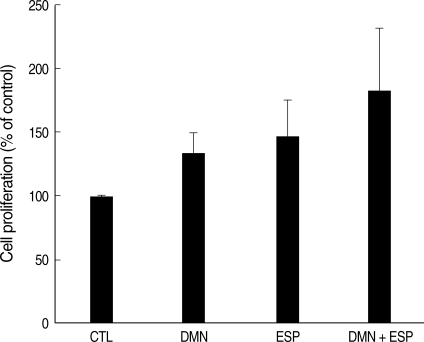
ESP and DMN synergistically affect the proliferation of HEK293T cells
To investigate the role of ESP and DMN in the proliferation of HEK293T cells XTT assay was performed. Cellular proliferation in HEK293T cells treated with ESP and/or DMN was increased compared to that in the controls (
Fig. 1). Although each compound increased proliferation when used alone, co-stimulation with DMN and ESP had the greatest effect on cell growth. The average increase compared to the control was: DMN, 132%; ESP, 152%; and DMN + ESP, 199% (
P = 0.0002). Thus, DMN and ESP synergistically affect cellular proliferation. All experiments were performed in triplicate.
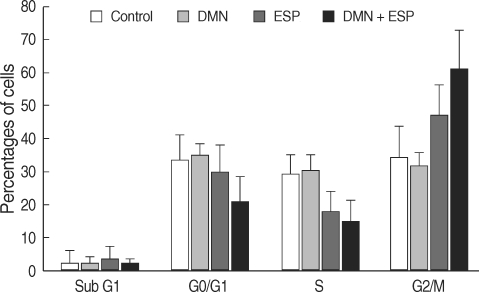
Cellular proliferation is tightly connected with cell cycle progression; thus, we monitored cell cycle progression using PI staining (
Fig. 2). In HEK293T cells treated with ESP, DMN, or ESP + DMN for 24 hr, the number of cells in the G0/G1 phase decreased. In contrast, the number of cells in the G2/M phase, which included cells undergoing rapid proliferation, was increased compared to the control (
Fig. 2). Fewer S-phase cells were identified in the ESP and ESP + DMN groups. The percentage of G2/M-phase cells in each group was: control, 33.8%; DMN, 32.1%; ESP, 50.1%; and ESP + DMN, 63.7% (
P = 0.0065).
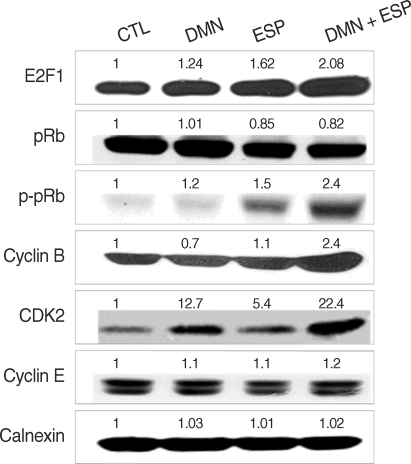
We used immunoblotting to detect the regulation of cell-cycle-related proteins in each group with calnexin as a loading control. The expression of several proteins, including E2F1, pRb, p-pRb, CDK2, and cyclin B, was upregulated, especially in the ESP + DMN group (
Fig. 3). In contrast, the p-pRb level was slightly decreased, implying that pRb degradation occurred. DMN alone induced CDK2 expression, whereas ESP enhanced the phosphorylation of pRb and slightly upregulated the level of CDK2. These results support the view that DMN and ESP synergistically affect cell cycle progression (
Fig. 2). As expected, the highest level of expression of cyclin B and CDK2 was detected in the DMN + ESP group. However, the level of CDK4 and cyclin E expression was similar in all of the groups (
Fig. 3).
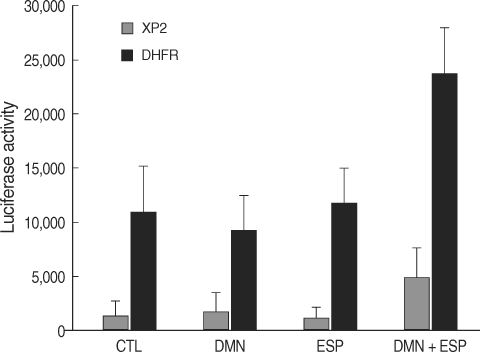
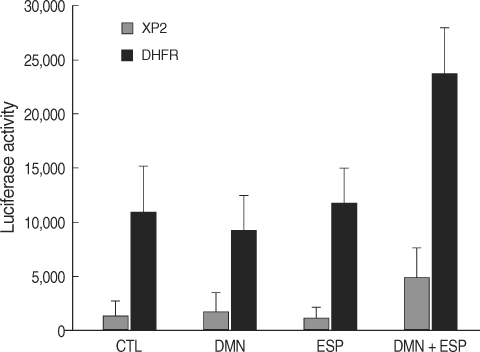
To determine whether the modulation of E2F1 activity is due to increased transcription of
E2F1, we performed transient transfection experiments using a luciferase reporter gene (
dhfr-luc) with pXP2-luc as a control [
25]. The gene
dhfr encodes an enzyme that catalyzes the synthesis of purines and thymidylate, which are necessary for DNA replication during S phase [
23,
24]. Furthermore,
dhfr contains E2F binding elements; thus, its expression is correlated with
E2F1 mRNA expression [
16,
25]. At 24 hr post-transfection, the HEK293T cells were plated and treated with ESP and/or DMN as indicated for an additional 24 hr. Luciferase activity was markedly increased in the transfected cells that were co-stimulated with ESP and DMN (
P = 0.014) (
Fig. 4).
DISCUSSION
Infection by
C. sinensis stimulates the bile duct epithelium, leading to the differentiation and proliferation of bile duct or periductal cells in infected individuals [
10]. Chronic inflammation induced by liver fluke infection is an important factor in cholangiocarcinogenesis, along with certain carcinogens, including nitrosamine compounds [
8,
26]. These compounds may be obtained from endogenous sources or from parasites [
7]. For example, NO synthesis is an important determinant of endogenous nitrosamine production in
O. viverrini-infected hamsters [
9]. In rats and hamsters infected with
C. sinensis, small ductular or periductal cells were the proliferation and differentiation [
6,
11,
12]. In addition, we reported that ESP from
C. sinensis induces the proliferation of HEK293T cells [
22]. In this study, we examined whether the carcinogen DMN has any influence on the proliferative effect of ESP in vitro.
Infection of
C. sinensis is regarded as an important, but preventable, cause of cancer [
27]. It has been more than 50 yr since cholangiocarcinogenesis was first associated with liver flukes [
27,
28]. Although co-treatment with DMN and metacercariae of
C. sinensis can induce cholangiocarcinoma in a hamster model [
6], the underlying mechanism is unknown. To understand how target cells respond to
C. sinensis antigens, we developed an in vitro system using the human epithelial cell line HEK293T. We stimulated HEK293T cells with ESP from
C. sinensis worms and/or the carcinogen DMN to provide a biological environment similar to that reported to cause cancer [
29]. We also focused on the proliferative activity in each group because uncontrolled proliferation is a key step in carcinogenesis.
As previously reported [
22], treatment with ESP increased the proliferation of the cells compared to the control level and was more effective than treatment with DMN (
Fig. 1). When HEK293T cells were co-stimulated with DMN and ESP, however, the degree of proliferation was increased further (
Fig. 1).
We also investigated the cell cycle distribution of the cells (
Fig. 2). Treatment with DMN did not result in a change in the cell cycle, whereas treatment with ESP decreased the proportion of cells in the G0/G1 and S phases and increased the proportion of G2/M-phase cells (
Fig. 2). Treatment with DMN + ESP maximized the proportion of G2/M-phase cells, implying that DMN and ESP synergistically affect cell cycle progression (
Fig. 2).
Cell cycle progression is driven by the coordinated regulation of activating CDKs and their positive regulatory cyclins. The
cdc2 and
cyclin B1 promoters are directly related to E2F [
20,
25]. Various studies have shown that E2F proteins influence the expression of genes that are involved in regulating the cell cycle at the G2/M phase, cell cycle progression, and DNA replication [
15,
21]. The role of E2Fs in controlling the transcription of
cdc2 and
cyclin B1 involves interactions with endogenous and G2/M-regulated promoters [
16,
18]. E2F1 is also a key factor in the development of cancer, given that several proto-oncogenes are regulated by E2Fs [
24,
27]. We therefore analyzed the expression of a number of cell-cycle-related proteins (
Fig. 3), including cyclin B1, which is an essential regulator of the G2/M transition [
15,
25]. Co-stimulation with ESP and DMN upregulated E2F1, cdK2, cyclin B1, and pRb phosphorylation, but the level of pRb was slightly reduced (
Fig. 3). Rb proteins physically interact with E2Fs, blocking their function as transcription factors and eventually causing cell cycle arrest. Once an Rb protein is phosphorylated, E2F molecules are free to move to the nucleus, where they function as transcription factors with DP [
16,
20]. The phosphorylation of pRb (
Fig. 3) implies that E2F1 may induce cell cycle progression, resulting in cellular proliferation, by acting as a transcription factor.
E2F1 expression is correlated with tumor proliferation, and the over-expression of E2F1 may produce very aggressive tumors with a high proliferation rate [
30]. In our system, co-stimulation with ESP and DMN induced the over-expression of E2F1 and down-regulation of pRb, indicating the involvement of the Rb-E2F1 pathway. The loss of Rb expression is consistent with Rb inactivation, and the hyperphosphorylation of pRb may release E2F1, reversing its growth-suppressive effect [
12,
15,
29].
The negative effect of strong E2F1 expression on Rb suppressor function may also be related to DNA damage and/or repair. ESP from
C. sinensis stimulates the proliferation of human embryonic kidney cells by increasing E2F1 expression [
29]. We hypothesized that the regulation of E2F1 is a key factor in the development of cancer in response to infection with liver fluke and exposure to a carcinogen such as DMN. We performed a
dhfr-promoter assay to determine whether the up-regulation of E2F1 occurs at the transcriptional level. Luciferase activity was markedly increased in cells co-stimulated with ESP and DMN (
Fig. 4).
In summary, the co-stimulation of human embryonic kidney cells (HEK293T) with the excretory/secretory products of C. sinensis and the carcinogen DMN induced cell proliferation via a synergistic effect on cell cycle progression. In particular, DMN up-regulated the expression of CDK2 while ESP induced the phosphorylation of pRB. Thus, treatment with DMN and ESP affects cellular proliferation, possibly initiating and promoting carcinogenesis. Although other changes may be involved in carcinogenesis, e.g., changes in the expression of cancer-related proteins, this hyper-proliferative effect may be the first step in the malignant transformation of normal cells.
ACKNOWLEDGEMENTS
This work was financially supported by grant numbers 800-20080583 from Korea Centers for Disease Control and Prevention and 04-2008-060-0 from the Seoul National University Hospital Research Fund.
References
Fig. 1Effects of ESP of C. sinensis and DMN on the proliferation of human embryonic kidney cells (HEK293T). Cellular proliferation in each treatment was determined using the XTT assay. Bars and whiskers indicate cell growth as the percentage of the control ± standard deviation (n = 4).

Fig. 2Effects of ESP of C. sinensis and DMN on the cell cycle distribution of HEK293T. Bars and whiskers indicate cell growth as the percentage of the control ± standard deviation (n = 4).

Fig. 3Expression of cell-cycle-related proteins in response to the ESP and DMN in HEK293T. Each sample was subjected to immunoblotting and probed with the indicated antibodies. Quantitative densitometry confirmed the observations presented in each blot and is shown as numbers.
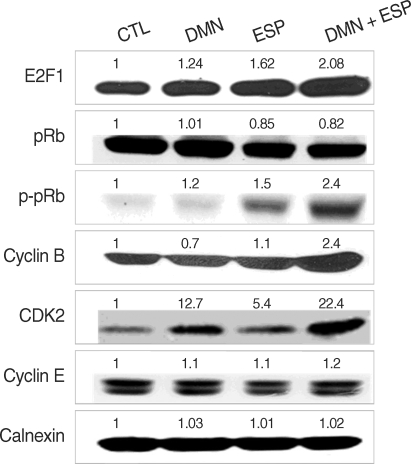
Fig. 4Treatment of ESP and DMN upregulated dhfr promoter-driven luciferase activity in HEK293T cells transfected with the luciferase reporter gene dhfr-luc (DHFR) or pXP2-luc (XP2, control). Bars and whiskers indicate the mean ± standard deviation (n = 3).