Abstract
Different functions have been attributed to CD4+CD25+Foxp3+ regulatory T-cells (Tregs) during malaria infection. Herein, we describe the disparity in Treg response and pro- and anti-inflammatory cytokines during infection with Plasmodium berghei ANKA between young (3-week-old) and middle-aged (8-month-old) C57BL/6 mice. Young mice were susceptible to cerebral malaria (CM), while the middle-aged mice were resistant to CM and succumbed to hyperparasitemia and severe anemia. The levels of pro-inflammatory cytokines, such as TNF-α, in young CM-susceptible mice were markedly higher than in middle-aged CM-resistant mice. An increased absolute number of Tregs 3-5 days post-inoculation, co-occurring with elevated IL-10 levels, was observed in middle-aged CM-resistant mice but not in young CM-susceptible mice. Our findings suggest that Treg proliferation might be associated with the suppression of excessive pro-inflammatory Th1 response during early malaria infection, leading to resistance to CM in the middle-aged mice, possibly in an IL-10-dependent manner.
-
Key words: Plasmodium berghei ANKA, cerebral malaria, CD4+CD25+Foxp3+ regulatory T cell, age, cytokine
INTRODUCTION
Cerebral malaria (CM) is one of the most severe complications of
Plasmodium infection and a major cause of death, primarily afflicting children aged 2-6 years in sub-Saharan Africa [
1]. CM appears to be mediated more by immunopathological host responses to infection than by the parasite per se [
2,
3]. Studies on malaria have progressively shown an important role for overwhelming the pro-inflammatory Th1 pathway in CM pathogenesis [
4,
5] and the subsequent combined effects of sequestration of parasitized red blood cells within blood vessels in the brain. However, the precise mechanism responsible for neuropathology remains unknown. Excessive serum levels of pro-inflammatory cytokines have been implicated in the pathogenesis of CM in murine models and human studies, with an association between higher mortality rate and elevated pro-inflammatory cytokine levels [
6,
7]. However, several have also revealed that pro-inflammatory cytokines are critical for the successful control and resolution of malaria infection in both humans and murine models [
8]. A weak pro-inflammatory response may lead to persistence and replication of parasites, while an excessive pro-inflammatory response may result in immunopathological consequences such as CM. Therefore, induction of an appropriate and effective immune response to malaria infection is needed for the host to subsequently control and eliminate this pathogen.
CD4
+CD25
+Foxp3
+ regulatory T cells (Tregs) play determinant roles in the preservation of self-tolerance and in the control of graft and tumor rejection and inflammation. Their abrogation leads to autoimmunity and inflammatory diseases in several experimental models [
9,
10]. Tregs also participate in the control of overwhelming responses to infectious agents such as viruses, bacteria, and protozoan parasites [
11,
12]. In malaria, Tregs expand during infection with the
Plasmodium berghei ANKA strain [
13,
14] and have been shown to inhibit the development of pathogenic Th1 cells, responsible for cerebral disease in resistant BALB/c mice [
13]. These results contrast with the detrimental effect associated with Tregs during
P. berghei ANKA infection in susceptible C57BL/6 mice [
14,
15]. In this infection model, depletion of Tregs results in a significant increase in survival, a minor but significant reduction in blood parasitemia, and an important reduction in parasite load in the brain and vasculature. A comparable delay in the onset of peak parasitemia has been reported during
P. berghei NK65 infection in mice depleted of Tregs [
16], and in the
P. yoelii 17XL infection model the elimination of Tregs allows BALB/c mice to control otherwise lethal infections [
17]. Moreover, during
Plasmodium falciparum infection in humans, the expansion of natural Tregs and the production of transforming growth factor-β (TGF-β) is correlated with higher parasite multiplication rates [
18,
19]. Altogether, these observations attribute contrasting functions to natural Tregs during
Plasmodium infections.
A study on age-related susceptibility and resistance to
P. berghei in mice revealed that 70% of 4-week-old C57BL/6 mice died from CM. However, only 20-30% of 10- and 16-week-old C57BL/6 mice developed CM [
20]. Our previous studies have shown that activation of Tregs is correlated with susceptibility in
P. yoelii 17XL-infected mice. Tregs can regulate the Th1 response by modifying dendritic cells whose expansion as well as increased IL-12 production following infection provide important co-stimulatory and cytokine signals to support the proliferation and activation of Th1 cells [
21]. Tregs mediate the incidence and outcome of CM in
P. berghei ANKA-infected mice by modifying the pro-inflammatory responses [
22-
24]. Herein, we compared the infection course and Treg response in young (3-week-old) and middle-aged (8-month-old) C57BL/6 mice infected with
P. berghei ANKA in order to elucidate the importance of Tregs in CM.
MATERIALS AND METHODS
Mice, parasites, and experimental infection
Female 3-week-old (young) and 8-month-old (middle-aged) C57BL/6 mice were purchased from Beijing Animal Institute (Beijing, China). P. berghei ANKA was kindly provided by Dr. Motomi Torii (Department of Molecular Parasitology, Ehime University Graduate School of Medicine, Ehime, Japan). Infections were initiated by intraperitoneal (i.p.) injection of 1×106
P. berghei ANKA parasitized erythrocytes for each group of young and middle-aged C57BL/6 mice. Parasitemia was monitored by counting the number of parasite-infected erythrocytes per 1,000 erythrocytes by light microscope examination of Giemsa-stained, thin (tail) blood smears. All experiments were performed in compliance with local Animal Ethics Committee requirements.
Spleen cell culture
Splenocyte culture was performed as previously described [
25]. Briefly, spleens from normal and infected mice were removed aseptically and pressed through a sterile fine-wire mesh with 10 ml RPMI 1640 supplemented with 10% heat-inactivated fetal calf serum (FCS), 25 mM HEPES, 0.12% gentamicin, and 2 mM glutamine. Cell suspensions were collected by centrifuging at 350
g for 10 min. Erythrocytes were lysed with cold 0.17 M NH
4Cl, and cells were washed twice with fresh medium. The viability of cells was determined by trypan blue exclusion and was always greater than 90%. Aliquots (500 µl/well) of cell suspensions (1×10
7/ml) were incubated in 24-well flat-bottom tissue culture plates (Falcon) in triplicate for 48 hr at 37℃ in a humidified 5% CO
2 incubator. Supernatant fractions were collected and stored at -80℃ until they were assayed.
A portion of the simultaneously infected mice was sacrificed at the indicated time to detect Tregs. Spleen cells from C57BL/6 mice were collected at different time points after infection. To assess Tregs, FITC-anti-CD4 and PE-anti-CD25 antibodies (clone PC61) were added to spleen cells and resuspended in 100 µl of PBS supplemented with 3% FCS for surface staining. The cells were fixed and permeabilized and intracytoplasmic staining was performed using APC-anti-Foxp3 (clone FJK16s) antibody. The cells were washed twice with PBS containing 1% FCS and resuspended in 300 µl of PBS. The cells were analyzed in a FACSCalibur cytofluorometer using CellQuest software. Viable cells were gated by forward and side scattering. Unless otherwise indicated, antibodies were purchased from BD Biosciences (San Jose, California, USA).
Detection of cytokines by ELISA
Levels of IFN-γ, TNF-α, and IL-10 were measured by commercial ELISA kits according to the manufacturer's protocol (R&D Systems, Minneapolis, Minnesota, USA). The OD values were read in a microplate reader at 450 nm. The concentration of cytokines in each sample was calculated using a standard curve generated using recombinant cytokines.
Statistical analysis
Data are presented as the mean±standard error of the mean (SE). Statistical significance of the differences was analyzed using the Student t-test or 1-way ANOVA (SPSS 17.0). A value of P<0.05 was considered significant.
RESULTS
Course of infection in young and middle-aged C57BL/6 mice
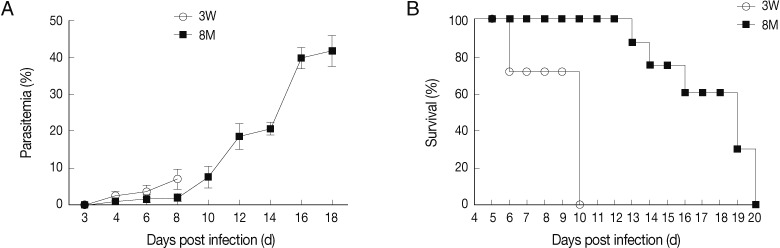
Young and middle-aged C57BL/6 mice infected with
P. berghei ANKA showed divergent courses of infection and disease severity. The young mice succumbed to
P. berghei ANKA-mediated CM on day 6 to 10 post-infection (PI) with characteristic signs of CM, including ruffled hair coat, rapid respiration, and lack of mobility, and parasitemia around 7%. Although the middle-aged mice died at day 13 to day 20 PI, the neurological signs of CM were not exhibited in this group, with a peak parasitemia of approximately 40% (
Fig. 1). These results indicated that the middle-aged C57BL/6 mice were resistant to CM during
P. berghei ANKA infection.
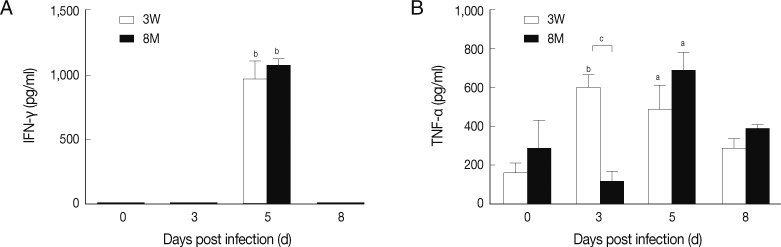
CM is hypothesized to result from a strong host pro-inflammatory response mediated by cytokines. Here, we determined the concentrations of IFN-γ and TNF-α in the supernatants of splenocyte cultures from young and middle-aged mice by ELISA. Infection led to significantly increased IFN-γ production on day 5 PI in both young and middle-aged mice (
P<0.01). The IFN-γ level was higher in middle-aged mice than in young mice, but the difference was not statistically significant (
Fig. 2A).
The change in TNF-α production by splenocytes differed between the young and middle-aged mice. TNF-α levels in young mice increased significantly on days 3 (
P<0.01) and day 5 PI (
P<0.05), whereas there was a significant elevation in middle-aged mice on day 5 PI only (
P<0.05). On day 3 PI, the TNF-α level in young mice was significantly higher than in middle-aged mice (
P<0.05,
Fig. 2B).
Our results showed that infection with malaria-causing parasites induced production of inflammatory cytokines, and the outcome of the infection was associated with the level and time of cytokine production.
Treg, CD4+ T cell, and IL-10 levels in young and middle-aged mice
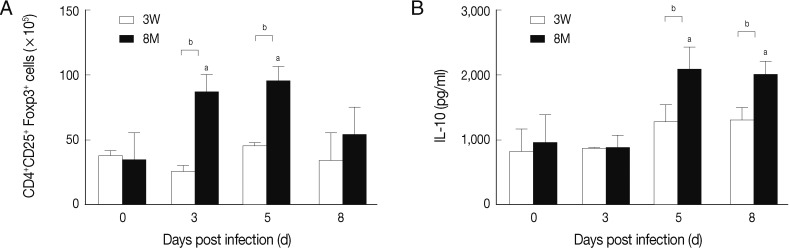
To compare the immunoregulatory effects of Tregs in young and middle-aged mice during malaria parasite infection, flow cytometric analysis was performed. By triple staining with FITC-anti-CD4, PE-anti-CD25, and APC-anti-Foxp3 monoclonal antibodies, Tregs in splenocytes from young and middle-aged mice infected with
P. berghei ANKA were evaluated. As shown in
Fig. 3A, there was no significant change in the number of Tregs in young mice at each time point compared with uninfected young mice, whereas in adult mice on day 3 PI there was a significant increase (
P<0.05) in the cell number up to 86.26×10
5 which then peaked at day 5 PI (
P<0.05) with an absolute cell number of 94.76×10
5.
To elucidate the potential mechanism of immunosuppression by Tregs, we evaluated the Th1 inhibitory cytokine, IL-10, in supernatants of splenocytes from parasite-infected mice. There was a slight but insignificant increase in the level of IL-10 in young mice after parasitic infection. In contrast, the IL-10 level in middle-aged mice increased significantly (
P<0.05) compared to both uninfected and young-infected mice and remained elevated from day 5 to day 8 PI (
Fig. 3B). Thus, our data reveal a marked difference in the kinetics of Treg proliferation and the production of immunoregulatory cytokine, IL-10, between young susceptible and middle-aged resistant mice during
P. berghei ANKA infection.
DISCUSSION
An appropriate immune response is critical in determining the outcome of malaria infection. Severe malaria, such as CM, is considered a complex multisystem disorder. The emergence of parasitemia and subsequent potent pro-inflammatory response are both essential for the occurrence of CM [
26], and the development of CM is associated with high levels of inflammatory cytokines during malaria infection [
27,
28]. A minimal pro-inflammatory response is beneficial in limiting parasitemia, but an exacerbated host response invariably leads to tissue damage. Neutralization of the pro-inflammatory cytokine IFN-γ in vivo is protective against murine CM and its associated mortality. IFN-γ
-/- [
29] and IFN-γ receptor (IFN-γR)
-/- [
30] mice are resistant to CM. Although IFN-γ is thought to be part of the immune system response to malaria infection, an immunopathological role is also quite conceivable [
31]. In humans, TNF-α is involved in the pathogenesis of human CM [
32].
P. berghei ANKA-infected mice present with higher cerebral levels of TNF-α [
33]. TNF-α is the principal cytokine mediator of CM in susceptible CBA/Ca mice [
6]. Consistent with these findings, our experimental results showed that the levels of IFN-γ and TNF-α increased markedly after malaria parasite infection in young CM-susceptible mice. In particular, the TNF-α level was significantly higher in young mice than in middle-aged CM-resistant mice on day 3 PI, which indicates that an early excessive pro-inflammatory Th1 response with high levels of IFN-γ and TNF-α might be responsible for the susceptibility of young C57BL/6 mice to CM.
Several groups of T-cells, particularly the immunosuppressive Tregs, have been shown to play a critical role in balancing protective immune responses and immune-mediated pathology. In our study, Tregs expanded in middle-aged CM-resistant mice during infection with
P. berghei ANKA as described previously [
13,
14], whereas in young CM-susceptible mice Tregs did not expand significantly. Thus, we demonstrated that this cell population has a regulatory role in the control of fatal pathogenesis. Moreover, the activation of Tregs accompanied by a high level of IL-10 was consistent with Tregs exerting their function in an IL-10-dependent manner [
34,
35]. IL-10 seems to have a host-protective role in murine malaria. In a susceptible mouse strain, administration of IL-10 gives some degree of protection against CM induced by
P. berghei ANKA, while in a resistant strain, a neutralizing anti-IL-10 antibody leads to a significant incidence of cerebral complications [
36]. In Vietnamese adults with severe malaria, plasma IL-10 levels were higher in those that died than in those who survived [
37], but CM victims have even lower levels. IL-10 is pleiotropic but a relevant mechanism could be its ability to inhibit the production of cytokines, such as TNF-α [
38], and it could also suppress excessive proinflammatory responses in human and experimental malaria [
39]. Thus, the expansion of Tregs and the high level of IL-10, which counteracts the production of TNF-α, might limit the excessive pro-inflammatory Th1 response in middle-aged mice, conferring resistance to CM, consistent with our previous study [
23]. However, in young mice, the low level of Tregs and IL-10 may not control the early excessive pro-inflammatory Th1 response, causing the young mice to die from CM.
In summary, the significant finding in this report is the importance of Treg response during infection with P. berghei ANKA in young CM-susceptible and middle-aged CM-resistant C57BL/6 mice. Our results raise the possibility that IL-10-dependent Tregs play an immunosuppressive role in the establishment of early excessive pro-inflammatory Th1 immunity, leading to resistance to CM in the middle-aged mice.
ACKNOWLEDGMENTS
We thank Dr. Motomi Torii (Ehime University Graduate School of Medicine, Ehime, Japan) for providing malaria parasite strains of P. berghei ANKA. We are also grateful to the Faculty of China Medical University, Department of Immunology for technical help and logistical support during the development of this work.
References
- 1. World Health Organization, Communicable Diseases Cluster. Severe falciparum malaria. Trans R Soc Trop Med Hyg 2000;94(suppl):S1-S90.
- 2. Hunt NH, Golenser J, Chan-Ling T, Parekh S, Rae C, Potter S, Medana IM, Miu J, Ball HJ. Immunopathogenesis of cerebral malaria. Int J Parasitol 2006;36:569-582.
- 3. Stevenson MM, Riley EM. Innate immunity to malaria. Nat Rev Immunol 2004;4:169-180.
- 4. Engwerda C, Belnoue E, Gruner AC, Renia L. Experimental models of cerebral malaria. Curr Top Microbiol Immunol 2005;297:103-143.
- 5. Ortolano F, Maffia P, Dever G, Hutchison S, Benson R, Millington OR, De Simoni MG, Bushell TJ, Garside P, Carswell HV, Brewer JM. Imaging T-cell movement in the brain during experimental cerebral malaria. Parasite Immunol 2009;31:147-150.
- 6. Grau GE, Fajardo LF, Piguet PF, Allet B, Lambert PH, Vassalli P. Tumor necrosis factor (cachectin) as an essential mediator in murine cerebral malaria. Science 1987;237:1210-1212.
- 7. Grau GE, Frei K, Piguet PF, Fontana A, Heremans H, Billiau A, Vassalli P, Lambert PH. Interleukin 6 production in experimental cerebral malaria: Modulation by anticytokine antibodies and possible role in hypergammaglobulinemia. J Exp Med 1990;172:1505-1508.
- 8. Riley EM, Wahl S, Perkins DJ, Schofield L. Regulating immunity to malaria. Parasite Immunol 2006;28:35-49.
- 9. Fontenot JD, Rasmussen JP, Williams LM, Dooley JL, Farr AG, Rudensky AY. Regulatory T cell lineage specification by the forkhead transcription factor Foxp3. Immunity 2005;22:329-341.
- 10. Schwartz RH. Natural regulatory T cells and self-tolerance. Nat Immunol 2005;6:327-330.
- 11. Belkaid Y, Sun CM, Bouladoux N. Parasites and immunoregulatory T cells. Curr Opin Immunol 2006;18:406-412.
- 12. Petersen PE. Global policy for improvement of oral health in the 21st century-implications to oral health research of world health assembly 2007, World Health Organization. Community Dent Oral Epidemiol 2009;37:1-8.
- 13. Nie CQ, Bernard NJ, Schofield L, Hansen DS. CD4+CD25+ regulatory T cells suppress CD4+ T-cell function and inhibit the development of Plasmodium berghei-specific Th1 responses involved in cerebral malaria pathogenesis. Infect Immun 2007;75:2275-2282.
- 14. Vigario AM, Gorgette O, Dujardin HC, Cruz T, Cazenave PA, Six A, Bandeira A, Pied S. Regulatory CD4+CD25+Foxp3+ T cells expand during experimental Plasmodium infection but do not prevent cerebral malaria. Int J Parasitol 2007;37:963-973.
- 15. Amante FH, Stanley AC, Randall LM, Zhou Y, Haque A, McSweeney K, Waters AP, Janse CJ, Good MF, Hill GR, Engwerda CR. A role for natural regulatory T cells in the pathogenesis of experimental cerebral malaria. Am J Pathol 2007;171:548-559.
- 16. Long TT, Nakazawa S, Onizuka S, Huaman MC, Kanbara H. Influence of CD4+CD25+ T cells on Plasmodium berghei NK65 infection in BALB/c mice. Int J Parasitol 2003;33:175-183.
- 17. Hisaeda H, Maekawa Y, Iwakawa D, Okada H, Himeno K, Kishihara K, Tsukumo S, Yasutomo K. Escape of malaria parasites from host immunity requires CD4+ CD25+ regulatory T cells. Nat Med 2004;10:29-30.
- 18. Walther M, Tongren JE, Andrews L, Korbel D, King E, Fletcher H, Andersen RF, Bejon P, Thompson F, Dunachie SJ, Edele F, de Souza JB, Sinden RE, Gilbert SC, Riley EM, Hill AV. Upregulation of TGF-β, Foxp3, and CD4+CD25+ regulatory T cells correlates with more rapid parasite growth in human malaria infection. Immunity 2005;23:287-296.
- 19. Edathodu J, Ali B, Alrajhi AA. CD4 validation for the World Health Organization classification and clinical staging of HIV/AIDS in a developing country. Int J Infect Dis 2009;13:243-246.
- 20. Pierrot C, Adam E, Lafitte S, Godin C, Dive D, Capron M, Khalife J. Age-related susceptibility and resistance to Plasmodium berghei in mice and rats. Exp Parasitol 2003;104:81-85.
- 21. Zheng W, Wang QH, Liu YJ, Liu J, Feng H, Wu JJ, Cao YM. Distinct host-related dendritic cell responses during the early stage of Plasmodium yoelii infection in susceptible and resistant mice. Parasite Immunol 2010;32:324-334.
- 22. Chen G, Liu J, Wang QH, Wu Y, Feng H, Zheng W, Guo SY, Li DM, Wang JC, Cao YM. Effects of CD4+CD25+Foxp3+ regulatory T cells on early Plasmodium yoelii 17XL infection in BALB/c mice. Parasitology 2009;136:1107-1120.
- 23. Wu JJ, Chen G, Liu J, Wang T, Zheng W, Cao YM. Natural regulatory T cells mediate the development of cerebral malaria by modifying the pro-inflammatory response. Parasitol Int 2010;59:232-241.
- 24. Zheng W, Wang QH, Feng H, Liu J, Meng HR, Cao YM. CD4+ CD25+Foxp3+ regulatory T cells prevent the development of Th1 immune response by inhibition of dendritic cell function during the early stage of Plasmodium yoelii infection in susceptible BALB/c mice. Folia Parasitol (Praha) 2009;56:242-250.
- 25. Greenberg J, Nadel EM, Coatney GR. The influence of strain, sex and age of mice on infection with Plasmodium berghei. J Infect Dis 1953;93:96-100.
- 26. Walther M, Jeffries D, Finney OC, Njie M, Ebonyi A, Deininger S, Lawrence E, Ngwa-Amambua A, Jayasooriya S, Cheeseman IH, Gomez-Escobar N, Okebe J, Conway DJ, Riley EM. Distinct roles for Foxp3 and Foxp3 CD4 T cells in regulating cellular immunity to uncomplicated and severe Plasmodium falciparum malaria. PLoS Pathog 2009;5:e1000364.
- 27. Edgerton C, Crispin JC, Moratz CM, Bettelli E, Oukka M, Simovic M, Zacharia A, Egan R, Chen J, Dalle Lucca JJ, Juang YT, Tsokos GC. IL-17 producing CD4+ T cells mediate accelerated ischemia/reperfusion-induced injury in autoimmunity-prone mice. Clin Immunol 2009;130:313-321.
- 28. Jain V, Armah HB, Tongren JE, Ned RM, Wilson NO, Crawford S, Joel PK, Singh MP, Nagpal AC, Dash AP, Udhayakumar V, Singh N, Stiles JK. Plasma IP-10, apoptotic and angiogenic factors associated with fatal cerebral malaria in india. Malar J 2008;7:83.
- 29. Yanez DM, Manning DD, Cooley AJ, Weidanz WP, van der Heyde HC. Participation of lymphocyte subpopulations in the pathogenesis of experimental murine cerebral malaria. J Immunol 1996;157:1620-1624.
- 30. Amani V, Vigario AM, Belnoue E, Marussig M, Fonseca L, Mazier D, Renia L. Involvement of ifn-gamma receptor-medicated signaling in pathology and anti-malarial immunity induced by Plasmodium berghei infection. Eur J Immunol 2000;30:1646-1655.
- 31. McGuirk P, Mills KH. Pathogen-specific regulatory T cells provoke a shift in the Th1/Th2 paradigm in immunity to infectious diseases. Trends Immunol 2002;23:450-455.
- 32. Brown H, Turner G, Rogerson S, Tembo M, Mwenechanya J, Molyneux M, Taylor T. Cytokine expression in the brain in human cerebral malaria. J Infect Dis 1999;180:1742-1746.
- 33. Adkins B, Leclerc C, Marshall-Clarke S. Neonatal adaptive immunity comes of age. Nat Rev Immunol 2004;4:553-564.
- 34. Belkaid Y, Piccirillo CA, Mendez S, Shevach EM, Sacks DL. Cd4+ cd25+ regulatory t cells control Leishmania major persistence and immunity. Nature 2002;420:502-507.
- 35. Brustoski K, Moller U, Kramer M, Hartgers FC, Kremsner PG, Krzych U, Luty AJ. Reduced cord blood immune effector-cell responsiveness mediated by CD4+ cells induced in utero as a consequence of placental Plasmodium falciparum infection. J Infect Dis 2006;193:146-154.
- 36. Kossodo S, Monso C, Juillard P, Velu T, Goldman M, Grau GE. Interleukin-10 modulates susceptibility in experimental cerebral malaria. Immunology 1997;91:536-540.
- 37. Day NP, Hien TT, Schollaardt T, Loc PP, Chuong LV, Chau TT, Mai NT, Phu NH, Sinh DX, White NJ, Ho M. The prognostic and pathophysiologic role of pro- and anti-inflammatory cytokines in severe malaria. J Infect Dis 1999;180:1288-1297.
- 38. Moore KW, de Waal Malefyt R, Coffman RL, O'Garra A. Interleukin-10 and the interleukin-10 receptor. Annu Rev Immunol 2001;19:683-765.
- 39. Lara C, Fayyad J, de Graaf R, Kessler RC, Aguilar-Gaxiola S, Angermeyer M, Demytteneare K, de Girolamo G, Haro JM, Jin R, Karam EG, Lepine JP, Mora ME, Ormel J, Posada-Villa J, Sampson N. Childhood predictors of adult attention-deficit/hyperactivity disorder: Results from the world health organization world mental health survey initiative. Biol Psychiatry 2009;65:46-54.
Fig. 1Parasitemia (A) and survival rate (B) of young and middle-aged mice infected with P. berghei ANKA. Percentages of parasitemia were calculated by counting the number of parasite-infected erythrocytes per 1,000 total erythrocytes by light microscope examination of Giemsa-stained, thin (tail) blood smears. Mortality was checked daily. Results are presented as the arithmetic mean of 9 mice per group±SE.
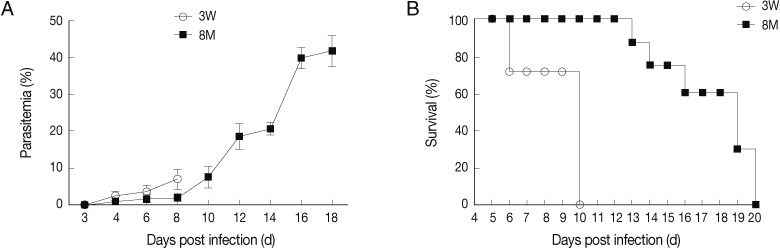
Fig. 2Level of IFN-γ and TNF-α in young and middle-aged mice with P. berghei ANKA infection. ELISA was performed to detect the level of IFN-γ and TNF-α in supernatants of cultured spleen cells from infected mice. Results are presented as the arithmetic mean of 9 mice per group±SE. aP<0.05, bP<0.01 versus corresponding values for uninfected control mice (day 0 PI). cP<0.05 indicates the comparison between young and middle-aged mice.

Fig. 3Treg number and IL-10 levels in young and middle-aged mice with P. berghei ANKA infection. The number of CD4+CD25+Foxp3+ Tregs in total spleen T cell population was measured by flow cytometry for C57BL/6 mice after inoculation. ELISA was performed to detect the level of IL-10 in supernatants of cultured spleen cells from infected mice. Results are presented as the arithmetic mean of 9 mice per group±SE. aP<0.05 versus corresponding values for uninfected control mice (day 0 PI). bP<0.05 indicates the comparison between young and middle-aged mice.