Abstract
The metacercariae of Centrocestus formosanus, a minute intestinal trematode of mammals and birds, were detected in the freshwater fish, Puntius brevis, from Vientiane Municipality, Lao PDR. The metacercariae were experimentally fed to mice, and adult flukes were recovered in their small intestines 7 days later. The adult flukes were morphologically characterized by having 32 (rarely 34) circumoral spines arranged in 2 alternative rows, a large bipartite seminal vesicle, an oval-shaped ovary, and an X-shaped excretory bladder. Based on these characters, the adults were identified as Centrocestus formosanus (Nishigori, 1924). The taxonomic significance of C. formosanus, in relation to a closely related species, C. caninus (Leiper, 1913), is briefly discussed. It has been first verified by adult worm recovery that C. formosanus is prevalent in Vientiane areas of Lao PDR, taking the freshwater fish, P. brevis, as a second intermediate host.
-
Key words: Centrocestus formosanus, Metacercaria, Adult, Heterophyid fluke, Fish
Centrocestus formosanus (Nishigori, 1924) (Digenea: Heterophyidae) is a minute food-borne intestinal trematode inhabiting the small intestine of birds and mammals, including chickens, ducklings, mice, rats, rabbits, dogs, cats, and foxes [
1]. The second intermediate hosts include various species of freshwater fishes, frogs, and toads in Taiwan, the Philippines, Hong Kong, and Vietnam [
1,
2]. This parasite can potentially infect humans, and an experimental human infection was reported successful in Taiwan [
3]. However, natural human infections with this fluke have never been documented. On the other hand,
Centrocestus caninus (Leiper, 1913), a closely related species to
C. formosanus, was reported from 2 human infections in Thailand [
4] and 3 humans in Lao PDR [
5].
Centrocestus formosanus is so far known to be distributed in Taiwan, China, Japan, the Philippines, Thailand, India, Hawaii, Vietnam, Croatia, USA, Mexico, and Colombia [
2,
6-
11]. In Lao PDR, metacercariae of
C. formosanus were reported once in a freshwater fish species,
Esomus longimana [
12], however, adult flukes have never been reported. The present study was performed to identify the adult flukes of
C. formosanus after experimental infection of the metacercariae to laboratory mice.
The metacercariae of C. formosanus were collected from the gill of Puntius brevis, a species of freshwater fish, collected in a local market of Vientiane Municipality, Lao PDR. Briefly, the fish body was divided into head with gills and body, minced, and artificially digested in an artificial gastric juice, made with 0.6% pepsin in 0.85% NaCl for 1 hr at 37℃ in a shaking water bath. The digested material was filtered through a sieve and rinsed several times with normal saline. Then, the metacercariae were isolated and counted under a stereomicroscope.
Male ICR mice, aged 4-6 wk, were purchased from an animal raising company (Dae-Han Biolink Co., Eumsung, Korea). The metercariae were fed orally to mice and killed on day 7 postinfection (PI). Their small intestines were resected, opened longitudinally in a Petri dish containing saline, and examined for worms using a stereomicroscope. The adult flukes were fixed in 10% neutral formalin under a cover slip pressure and stained with Semichon's acetocarmine, cleared in xylene, and mounted in Canada balsam. The stained specimens were microscopically examined, and measured. All measurements are given in micrometers, unless otherwise stated. Several adult specimens collected from the small intestines of mice at day 7 PI were prepared for scanning electron microscopy as described previously [
13].
Forty-two fish,
P. brevis, were examined, and a total of 490 metacercariae of
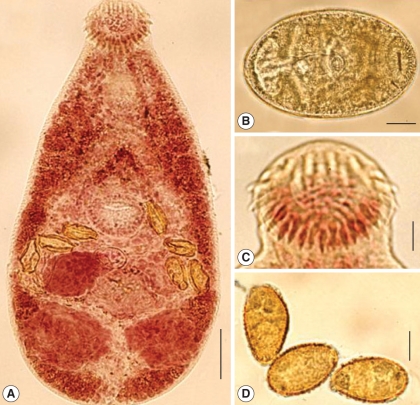
C. formosanus were harvested from the head with gills. The mean metacercarial density was 11.7 per fish. The encysted metacercariae were ellipsoidal, 178 ± 15 µm in length and 143 ± 5 µm in width (n = 10), and yellowish brown. The oral sucker was located at the anterior terminal. The circumoral spines were arranged in 2 alternative rows around the oral sucker, totaling 32 in number. The ventral sucker, about a half size of the oral sucker, was located in the middle portion of the body (
Fig. 1A, B). The metacercariae of
C. formosanus were grouped and infected orally to 5 mice. At day 7 PI, a total of 46 adult flukes (worm recovery rate; 9.4%) were recovered from the small intestine. The adult worms were identified as
C. formosanus, based on the following morphological features and measurements (n = 10) (
Table 1).
Morphology of adult worms (Figs. 1, 2)
Body ovoidal with broad posterior half (
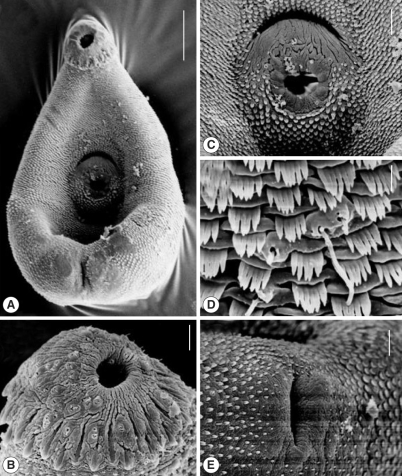
Fig. 1A), covered with scale-like tegumental spines (
Fig. 2A) from anterior to posterior ends. Oral sucker terminal, and 32 (34 in 1 of 10 specimens) circumoral spines arranged in 2 alternative rows (
Figs. 1C,
2B). Esophagus short. Ceca large, bifurcated about midway between ventral and oral suckers, terminated slightly in front of ovary. Ventral sucker smaller than oral sucker and located at median 2 / 5 of body. Round non-ciliated and uni-ciliated papillae and tegumental spines located on half of lower part of ventral sucker but upper part of ventral sucker, between ventral sucker and genital pore, without tegumental spines (
Fig. 2C). In mid-dorsal tegument, spines pectinate, with long uniciliated sensory papillae (
Fig. 2D). Seminal vesicle large, bipartite, with anterior part transversely elongated and posterior one rounded. Genital pore anterior to ventral sucker. Ovary oval, located on right side of posterior half of body. Seminal receptacle lies in middle of posterior half of body, dorsal to ovary, between seminal vesicle and anterior border of excretory bladder. Laurer's canal prominent, opening dorsally. Mehlis' gland and ootype opposite to ovary. Uterus short, coiled between ovary and seminal vesicle, filled with 4-16 (av. 10.4) eggs. Vitelline follicles large, scattered laterally from posterior end to posterior border of pharynx. Eggs oval, shell surface with lattice design, yellowish brown, operculum distinct (
Fig. 1D). Excretory bladder X-shaped, with short posterior tube. Excretory pore terminal, with low density of tegumental spines (
Fig. 2E).
In the present study, the adult flukes of
C. formosanus have been first identified in Lao PDR. The worm size was slightly smaller than those reported by previous authors [
1,
3], but it is considered to have been due to a shorter developmental period in our specimens. The important morphological features included the presence of 32 (rarely 34) circumoral spines arranged in two alternative rows, a large bipartite seminal vesicle, an oval-shaped ovary, egg shell surface with lattice design, and an X-shaped excretory bladder. The freshwater fish,
Puntius brevis, is suggested to be a source of animal or, possibly human, infections with
C. formosanus.
In the speciation of flukes belonging to the genus
Centrocestus, the number and size of circumoral spines, the margin forms of the ovary and testes, the size and number of eggs in the uterus, the ridge patterns on the egg shell surface, the level of ceca ends, the shape of excretory bladder, and the presence of prepharynx were adopted as useful keys [
1,
4]. Among these characters, the number of circumoral spines was accepted as the most reliable one, although the number varied showing a range in some species, at times overlapping one another [
1,
4]. Counting the exact number is difficult in some specimens. Waikagul et al. [
4] classified the reported species of Centrocestus into 3 big groups, i.e., those with 26-36 spines (
C. yokogawai,
C. caninus,
C. longus,
C. formosanus,
C. cuspidatus, and
C. asadai), 38-48 spines (
C. nycticoracis,
C. kurokawai, and
C. armatus), and 50-60 spines (
C. polyspinosus). Among the first group, they finally recognized only 2 species,
C. cuspidatus, which has a V-shaped excretory bladder, and
C. caninus, which has an X-shaped excretory bladder [
4]. Species of the first group other than the two, i.e.,
C. yokogawai,
C. longus,
C. formosanus, and
C. asadai, were regarded similar and synonymized with
C. caninus [
4].
However, the synonymy of
C. formosanus with
C. caninus is put to a question because of several reasons. First, the number of circumoral spines is 26-30 in
C. caninus [
4,
14], whereas that of
C. formosanus is 30-36 [
1,
3,
15], not overlapping significantly. The latter has in most cases 32 spines [
1,
3,
15] and rarely 30, 34, or 36 [
1]. Therefore, the number of circumoral spines may be a unique feature for both
C. formosanus and
C. caninus. Second,
C. caninus was originally described based on only 1 specimen obtained from a dog in Taiwan [
14]. The number of circumoral spines was counted 28, but, as the preservation status of the worm was not good, it was suggested that some of the spines originally present may have been lost [
14,
16,
17]. The worm was tentatively designated as a variant of
C. cuspidatus, i.e.,
C. cuspidatus caninus, until material for further study would be available [
14]. Separately from this,
C. formosanus was found and described as a new species in Taiwan [
3], and Faust and Nishigori [
17] considered that Leiper's specimen of
C. cuspidatus caninus was in fact
C. formosanus. Yamaguti [
18] raised
C. cuspidatus caninus up to a specific level, i.e.,
C. caninus, and synonymized
C. formosanus,
C. longus, and
C. yokogawai with
C. caninus without appropriate reasons. Later, he retained
C. formosanus as a valid species, whereas no more describing
C. caninus [
19]. A third point is that
C. caninus was not reported for a long time after it was first reported based on only 1 specimen in 1913 [
14], until 1997 when 6 specimens recovered from 2 human infections in Thailand were assigned as
C. caninus [
4]. We believe that treating
C. caninus as a specific level [
18] is problematic, and even if
C. caninus (as
C. cuspidatus caninus) had in fact more than 28 circumoral spines, for example, 32,
C. formosanus should have the taxonomic priority.
Among the
Centrocestus species, those reported from human infections are
C. armatus (experimental and natural) [
20,
21],
C. kurokawai (natural) [
22],
C. cuspidatus (natural) [
23],
C. caninus (natural) [
4,
5], and
C. formosanus (experimental) [
3]. In the report of human infections from Lao PDR [
5], the worm was tentatively designated, because the number of specimens was only 4 and detailed morphological studies were not possible. With regard to
C. formosanus, the possible presence of natural human infections has been suggested [
15,
24]. In this regard, epidemiological studies are required to verify this suggestion.
The second intermediate hosts of
C. formosanus were reported to be a variety of freshwater fishes,
Carassius auratus,
Pseudorasbora parva,
Channa formosana,
Gambusia affinis,
Ophicephalus tadianus,
Misgurnus anguillicaudatus,
Zacco platypus,
Cyprinus carpio,
Glossogobius giurus,
Anabas testudineus, and
Puntius semifasciolatus, frogs,
Rana limnocharis, and toads,
Bufo melanostictus [
1]. The present study adds
P. brevis as a new second intermediate host of
C. formosanus. The metacercariae are encysted mostly in the gills of fish [
1,
7], but in heavily infected fish a small number of metacercariae may also be present under the scale of muscle [
1]. As the gills are generally discarded during cooking fishes, infected people may be few in number. However, caution should be paid as the number of uterine eggs in
Centrocestus spp. adults is small, less than 30-40 eggs per worm [
1,
4,
25], and the chance of detecting eggs in fecal examination is low particularly in light infection cases. Moreover, the eggs of
C. formosanus closely resemble those of
O. viverrini, having lattice patterns on the egg shell; hence, researchers may find difficulty in differentiating them in fecal examinations.
References
Fig. 1The metacercaria and adult of Centrocestus formosanus. (A) An adult worm recovered from an ICR mouse 7 days after infection. Stained with acetocarmine (Bar: 40 µm). (B) A metacercaria from the freshwater fish, Puntius brevis, from Lao PDR (Bar = 30 µm). (C) Oral sucker of an adult worm with circumoral spines (Bar = 100 µm). (D) Eggs showing surface lattice patterns (Bar = 14 µm).

Fig. 2Scanning electron microscopic views of Centrocestus formosanus recovered from Lao PDR. (A) Whole ventral view of an adult worm (Bar = 40 µm). (B) Oral sucker with circumoral spines (Bar = 60 µm). (C) Ventral sucker and genital pore (Bar = 10 µm). (D) Tegumental spines and papillae in mid-dorsal region (Bar = 1 µm). (E) Tegument near the excretory pore (Bar = 4 µm).

Table 1.Comparative measurements
a of
Centrocestus formosanus by different authors
Table 1.
|
Present study |
Nishigori (1924) |
Chen (1942) |
|
Body length × width |
286 (245-325) × 192 (155-220) |
333 (289-389) × 201 (187-238) |
461 × 212 |
|
No. of oral spine |
32 (32-34) |
32 |
30-36 |
|
Oral sucker |
52 (45-58) × 43 (38-50) |
50 × 47 |
60 × 56 |
|
Ventral sucker |
48 (45-55) × 35 (33-45) |
39-55 × 36-43 |
52 × 55 |
|
Pharynx |
32 (28-34) × 26 (20-30) |
40-47 × 20-35 |
41 × 42 |
|
Ovary |
60 (50-80) × 42 (34-46) |
54 (50-63) × 40 (37-43) |
83 × 67 |
|
Right testis |
65 (45-93) × 38 (24-50) |
89 (77-100) × 43 (37-58) |
106 × 71 |
|
Left testis |
66 (55-88) × 40 (30-63) |
78 (55-100) × 42 (35-55) |
101 × 74 |
|
Anterior seminal vesicle |
81 (63-93) × 18 (14-23) |
122 (80-112) × 31 (15-33) |
78 × 35 |
|
Posterior seminal vesicle |
41 (30-45) × 28 (24-43) |
81 (63-83) × 26 (15-38) |
69 × 37 |
|
Seminal receptacle |
46 (25-60) × 37 (23-43) |
53 (43-63) × 50 (43-60) |
80 × 62 |
|
Eggs |
34 (30-36) × 17.1 (15-19) |
33 (33-35) × 17 (17-20) |
33 × 16 |