Abstract
Neospora caninum (Apicomplexa; Sarcocystidae) is a protozoan that causes abortion in cattle, horses, sheep, and dogs as well as neurological and dermatological diseases in dogs. In the central nervous system of dogs infected with N. caninum, cysts were detected that exhibited gliosis and meningitis. Flavonoids are polyphenolic compounds that exhibit antibacterial, antiparasitic, antifungal, and antiviral properties. In this study, we investigated the effects of flavonoids in a well-established in vitro model of N. caninum infection in glial cell cultures. Glial cells were treated individually with 10 different flavonoids, and a subset of cultures was also infected with the NC-1 strain of N. caninum. All of the flavonoids tested induced an increase in the metabolism of glial cells and many of them increased nitrite levels in cultures infected with NC-1 compared to controls and uninfected cultures. Among the flavonoids tested, 3',4'-dihydroxyflavone, 3',4',5,7-tetrahydroxyflavone (luteolin), and 3,3',4',5,6-pentahydroxyflavone (quercetin), also inhibited parasitophorous vacuole formation. Taken together, our findings show that flavonoids modulate glial cell responses, increase NO secretion, and interfere with N. caninum infection and proliferation.
-
Key words: Neospora caninum, glia, flavonoid, nitric oxide (NO)
INTRODUCTION
Neospora caninum (Apicomplexa; Sarcocystidae) is a protozoan that mainly causes dermatological and neurological disorders for which the dog is the definitive host [
1]. In cattle, the intermediate host of this parasite,
N. caninum causes a range of economic losses that are associated with abortion and a decline in milk production [
2].
N. caninum was first isolated in 1988 from the tissues of a naturally infected dog [
3,
4]. Since then, new strains have been isolated, such as NC-1 [
5,
6].
To elucidate the pathogenesis of infection with this parasite in the central nervous system (CNS), our group developed an in vitro model of
N. caninum infection in primary glial cell cultures [
7]. In this model, it was observed that astrocytes and microglia responded to infection by producing proinflammatory and regulatory cytokines, i.e., tumor necrosis factors-α (TNF-α) and IL-10, as well as the neurotoxic free radical nitric oxide (NO) [
7,
8,
9]. These results show that glial cells are targets of
N. caninum and that astrocytes may contribute to the CNS immune responses to the parasites.
Flavonoids are naturally occurring polyphenolic compounds that are present in a variety of fruits, vegetables, cereals, teas, wines, and fruit juices. They are secondary metabolites formed in plants from the aromatic amino acids phenylalanine and tyrosine as well as from malonate [
10]. In Brazil and several other countries, antibacterial, antiparasitic, antifungal, and antiviral properties have been attributed to flavonoids [
11,
12,
13]. Studies have shown that the flavonoid quercetin is able to inhibit the growth of
Leishmania danovani [
14],
L. panamensis [
15], and
L. amazonensis [
16]. Apigenin and the biflavone fukugetin also displayed considerable inhibition of
Trypanosoma cruzi [
17,
18]. The isoflavone genistein efficiently blocked host cell egress of
Toxoplasma gondii by more than 50% [
19].
Flavonoids have been shown to modulate glial cell responses [
20]. Recent findings from our group showed that flavonoid rutin induces activation of astrocytes and microglia and production of TNF-α and NO [
21]. Thus, in the present study, we evaluated whether the treatment with flavonoids induces glial activation and interferes with
N. caninum infection and proliferation.
MATERIALS AND METHODS
All of the experimental procedures were performed in accordance with the standards of the Ethics Committee on Animal Care of Bahia Federal University, Brazil.
Glial cell cultures
Mixed primary cultures of astrocytes and microglia were prepared according to the method of Cookson et al. [
22] and modified by Silva et al. [
21]. Newborn Wistar rats were decapitated on postnatal day 0-2, and their cerebral cortices were removed. After mechanical dissociation, the obtained cells were cultured in Dulbecco's modified Eagle's medium (DMEM, Cultilab, Campinas, São Paulo, Brazil) supplemented with 6.25 g/ml gentamicin, 2 mM L-glutamine, 0.011 g/L pyruvate and 10% foetal calf serum (Cultilab, Campinas, São Paulo, Brazil). The cells were seeded in 100 mm polystyrene culture plates (TPP) at a density of 3×10
6 cells/plate. After 2-3 weeks in culture, the cells were trypsinized and replated in microtiter dishes with 40 mm, 24 or 96 wells, depending on the experiment, at a density of 1×10
4 cells/cm
2.
Vero cells (from African green monkey kidneys) were maintained in DMEM enriched with 5% fetal bovine serum at 37℃ under 5% CO
2, with regular changes every 48 hr. After the formation of a confluent cell monolayer, the Vero cells were infected with tachyzoites of
N. caninum strain NC-1 [
3,
4], which was obtained from the Laboratory of Parasitology at the Veterinary Medicine Hospital of UFBA, Brazil. Glial cell primary cultures pretreated with flavonoids for 24 hr were infected with
N. caninum tachyzoites according to the protocol described by Pinheiro et al. [
7,
8]. Briefly, at the time of infection, the Vero cells were mechanically injured using a sterile mini-rod, and the parasites were purified by exclusion chromatography in a Sephadex G25 column (Sigma, St. Louis, Missouri, USA). The number of parasites was counted, and a parasite/cell ratio of 1:1 was adopted for glial cell infection.
Glial cells were treated individually with 10 different flavonoids: 7-dimethoxyflavone (F1); 7,8-dihydroxyflavone (F2); 3',4'-dihydroxyflavone (F3); chrysin, 5,7-dihydroxyflavone (F4); apigenin, 4',5,7-trihydroxyflavone (F5); luteolin, 3',4',5,7-tetrahydroxyflavone (F6); kaempferol, 3,4',5,7-tetrahyroxyflavone (F7); fisetin, 5-deoxyquercetin, 3,3',4',7-tetrahydroxyflavone (F8); quercetin, 3,3',4',5,6-pentahydroxyflavone (F9); or rutin, 3 ramnoglicoside of 3,3',4',5,6-pentahydroxyflavon (F10). These compounds were dissolved in dimethylsulfoxide (DMSO) and diluted in culture medium to a final concentration of 50 µM.
To study the effect of flavonoids in uninfected conditions, glial cells were treated with each flavonoid separately for 24 or 48 hr. To investigate whether pretreatment with flavonoids interferes with NC-1 infection, another group of glial cell cultures were treated with flavonoids 24 hr before infection with N. caninum and analyzed at 24 hr post-infection, representing 48 hr of treatment with flavonoids. Controls received only DMSO (0.1%) at the same dilution used in the treatment of cultures with flavonoids. Negative controls were run without treatment and/or infection. Four samples for each experimental variable were performed and repeated in 3 independent experiments.
Evaluation of N. caninum proliferation
At 24 hr after infection with N. caninum, glial cells were disrupted mechanically with a mini-rod followed by resuspension using a 5 ml syringe and a 0.21 G needle. Counts of intracellular and extracellular NC-1 tachyzoites were performed using a Neubauer chamber under an optical phase microscope (Nikon TS-100, Tokyo, Japan) in 2 samples for each flavonoid experimental variable and were repeated by 3 independent experiments. The results are presented as the number of tachyzoites/µl, adjusted to a final volume of 300 µl.
Glial cell viability assay
The viability of glial cells was quantified by measuring the activity of mitochondrial dehydrogenases using the 3-(4,5-dimethylthiazol-2-yl)-2,5-difeniltetrazolium bromide (MTT) assay [
23]. After treatment, the cultures were incubated with MTT at a final concentration of 1 mg/ml for 2 hr. Thereafter, the cells were lysed using 20% (w/v) sodium dodecyl sulphate (SDS) and 50% (v/v) dimethyl formamide (DMF) (pH 4.7) and were maintained overnight at 37℃ to dissolve formazan crystals. Absorbance was measured at 540 nm. Three independent experiments were conducted with 4 replicate wells for each experimental variable. The mean absorbance of the control group (0.1% DMSO) was considered to be 100% cell viability. There was no significant difference between the controls with DMSO and the negative controls without DMSO.
To identify
N. caninum tachyzoites, fixed cultures were rehydrated with PBS for 30 min at room temperature. First, endogenous peroxidase activity was blocked for 10 min with 3% H
2O
2 in PBS. Non-specific binding sites were blocked with 0.1% fetal calf serum for 30 min. Primary glial cells cultivated on glass coverslips were rinsed 3 times in PBS and incubated with a primary goat polyclonal anti-NC antibody (1:500 in PBS overnight) (P081121-006 VMRD, Pullman, Washington, USA). The cells were then incubated with a peroxidase-conjugated goat anti-mouse IgG antibody (1:1,000 in PBS for 1 hr) (Sigma, St. Louis, Missouri, USA). Tachyzoites were labelled with brown coloring using a 0.3% 4-Cl-α-naphthol/methanol solution diluted in PBS (1:5) containing H
2O
2 (0.33 µl/ml) and incubated at room temperature for 30 min. To identify glia cell morphology, co-staining was performed using the protocol established by Rosenfeld [
24], as described by Silva et al. [
21]. The number of cells and immunoreactive parasitophorous vacuoles containing
N. caninum tachyzoites were determined under the microscope at ×20 magnification in a 0.29 mm2 field, and the percentage of parasitophorous vacuoles in relation to the total number of cells in the field was determined for each experimental condition.
The production of NO was assessed as nitrite (NO
2) accumulation in the culture medium using a colorimetric test based on Griess reagent [
25], according to the protocol modified by Jesus et al. [
26].
Statistical analyses were performed using Graph Pad Prism 5.0 software (San Diego, California, USA). Values from different treatments were analyzed with a 1-way ANOVA followed by a Student-Newman-Keuls post-test to determine differences between groups for individual parameters. Student's t-test was used to compare 2 different groups of flavonoids at the same concentration. To compare nonparametric data, the Mann-Whitney test was used. The results are presented as the mean±SD. All statistical tests were considered significant at P<0.05.
RESULTS
Effect of flavonoids on viability of glial cells infected with N. caninum
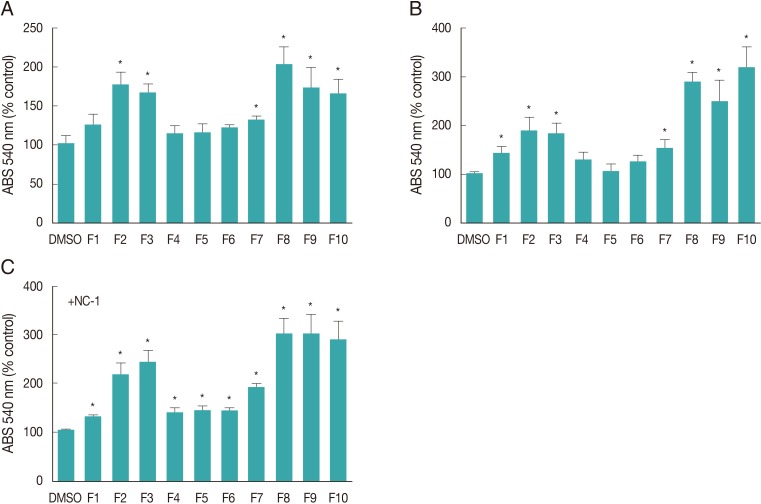
The toxicity of flavonoids F1 through F10 in glial cells was determined using the MTT assay. We observed that compared with control cultures (0.1% DMSO), flavonoids F2, F3, F7, F8, F9, and F10 (50 µM) induced an increase in cell viability in uninfected glial cell cultures (
Fig. 1A, B) after 24 hr and 48 hr of treatment. Additionally, an increase in cell viability was observed in NC-1 infected cultures pretreated with all flavonoids (
Fig. 1C). These effects were more pronounced in the cultures treated with flavonoids F2, F3, F8, F9, and F10.
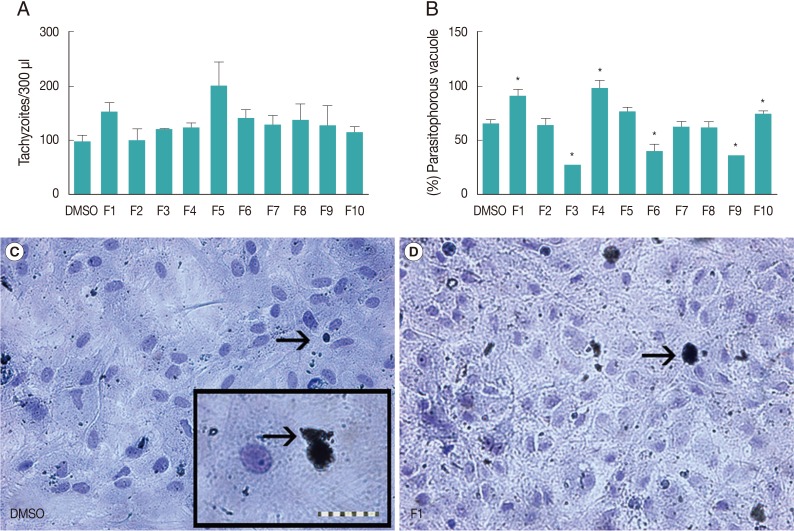
To investigate whether a 24 hr pretreatment of glial cells with flavonoids interferes with NC-1 infection, we quantified the number of tachyzoites and the proportion of glial cells with parasitophorous vacuoles in infected cultures with or without flavonoid pretreatment.
Fig. 2A shows the numbers of NC-1 tachyzoites present in cell lysates 24 hr after infection. The numbers of tachyzoites in cultures pretreated with flavonoids did not differ from those in cultures performed under control conditions (DMSO 0.1%). This may be because dead and alive tachyzoites did not differ morphologically, as observed under an optical phase microscope, and were counted together. However, analysis of NC-1 by immunocytochemistry and Rosenfeld staining showed that flavonoid pretreatment altered the proportion of glial cells presenting parasitophorous vacuoles. As shown in
Fig. 2, approximately 64.4% (±9.2% SD) of the cells in cultures infected with NC-1 under control conditions (DMSO 0.1%) exhibited parasitophorous vacuoles. Additionally, F3, F6, and F9 significantly reduced the proportion of cells with vacuoles. However, flavonoids F1, F4, and F10 increased the proportion of cells infected with NC-1, and this difference was statistically significant.
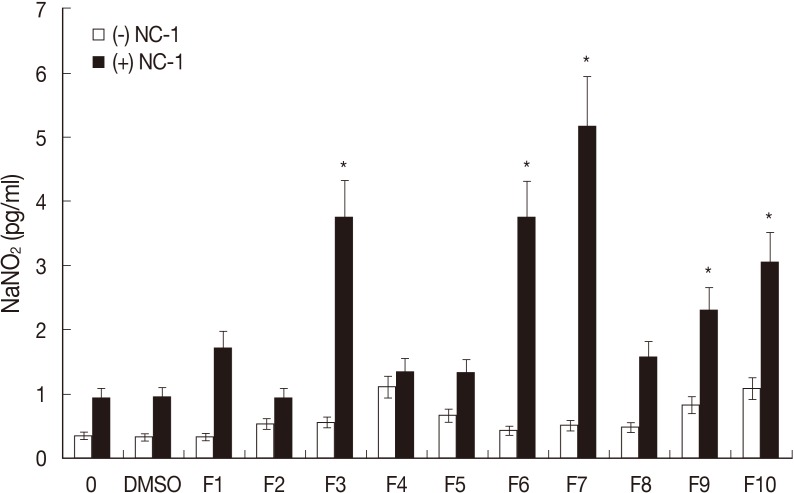
To investigate whether flavonoids and NC-1 infection interfere with glial cell functioning, the release of the trophic factor NO was evaluated. Nitrite levels in the culture media of control cultures (0.1% DMSO), from which NO levels were inferred, were very low (0.96 pg/ml) (
Fig. 3), and no significant increase in nitrite levels was observed following the 50 µM flavonoid treatments. However, an increase in nitrite levels was observed in cultures exposed to flavonoids F3, F6, F7, F9, and F10 and infected with NC-1, with these levels reaching 3.75, 3.72, 5.17, 2.31, and 3.05 pg/ml, respectively.
DISCUSSION
Among the tested flavonoids, F1, F2, F3, F7, F8, F9, and F10 induced a significant increase in MTT metabolism in both glial cell cultures infected with NC-1 and uninfected cultures. MTT conversion primarily occurs via mitochondrial dehydrogenases and is dependent on respiratory chain activity [
23]. Furthermore, increased activity of mitochondrial dehydrogenases in culture systems can be attributed either to an overall increase in metabolic activity or an increase in the number of MTT-metabolizing cells due to cell proliferation. Thus, it cannot be ruled out that flavonoids F1, F2, F3, F7, F8, F9, and F10 may have induced an increase in the metabolic activity of glial cells, particularly astrocytes, which are associated with the induction of microglial proliferation. Flavonoids F4, F5, and F6 all contain hydroxyl groups at positions 5 and 7 of the A ring, which may be linked to the biological response patterns observed.
Another pattern of glial cell responses resulting from flavonoid treatment and NC-1 infection was related to the proportion of cells with parasitophorous vacuoles. The biology of
N. caninum is characterized by rapid infection and spreading in the cytoplasm of host cells. In glial cell cultures, it has been determined that these parasites replicate in approximately 6 hr [
7,
8]. Apparently, pretreatment of glial cells with flavonoids F2, F5, F7, F8, and F10 did not affect the proliferation of this parasite. The cultures pretreated with flavonoids showed a proportion of parasitophorous vacuoles similar to that observed under control conditions. However, pretreatment of glial cells with flavonoids F3, F6, and F9 clearly reduced the proportion of infected cells presenting parasitophorous vacuoles containing NC-1. These molecules all present hydroxyls groups at the 3' and 4' positions of the B ring of their structures, forming a catechol nucleus, which has been attributed to toxicity in different biological systems. This characteristic may confer a toxic effect of flavonoids on NC-1 if this structure remains intact and biologically active at the time of infection. Moreover, this result may be due to an indirect effect of flavonoids on glial cell responses that interfered with the infection and multiplication of parasites in these host cells. In the mammalian CNS, NO can be produced via a free radical generation system. This system is absent in glial cells, but NO production can be stimulated through the conversion of arginine to citrulline via inducible NO synthase. Expression of this enzyme has been detected in cultures of glial cells and astrocytes in neuropathological samples [
27]. In the present study, a significant increase in the accumulation of nitrite, the stable form of NO, in the culture medium of cells treated with flavonoids was observed. However, this induction of NO production was observed in cultures infected with NC-1 when they were pretreated with flavonoids F3, F6, F7, F9, and F10. These findings suggest that there was a relationship between NO production and the prevention of NC-1 infection in the glial cell cultures.
Taken together, our findings clearly show that glial cells (astrocytes and microglia) respond differently to flavonoids exhibiting distinct degrees of hydroxylation and that these compounds induce glia activation, which interferes with N. caninum infection. Among the flavonoids tested and the evaluation parameters adopted in this study, F3 (3',4'-dihydroxyflavone), F6 (luteolin, 3',4',5',7-tetrahydroxyflavone), and F9 (quercetin, 3,3',4',5,6-pentahydroxyflavone) presented the most promising anti-parasitic potential. These drugs both induced glial cell activation and secretion of NO and inhibited N. caninum parasitophorous vacuole formation.
Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq)Fundação de Amparo à Pesquisa e Extensão do Estado da Bahia (FAPESB)
Notes
-
We have no conflict of interest related to this work.
ACKNOWLEDGMENTS
This work was supported in part by grants from the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) and the Fundação de Amparo à Pesquisa e Extensão do Estado da Bahia (FAPESB), Brazil. We gratefully acknowledge the research support provided by the Programa de Pos-graduação em Ciência Animal dos Trópicos/UFBA, Brazil. We also gratefully acknowledge Dr. Guy G. Chabot, Laboratoire de Pharmacologie Chimique et Génétique, Université Paris Descartes, France for kindly providing the flavonoids used in this study.
References
- 1. McAllister MM, Dubey JP, Lindsay DS, Jolley WR, Wills RA, McGuire AM. Dogs are definitive hosts of Neospora caninum. Int J Parasitol 1998;28:1473-1478.
- 2. Anderson ML, Blanchard PC, Barr BC, Dubey JP, Hoffman RL, Conrad PA. Neospora-like protozoan infection as a major cause of abortion in California dairy cattle. J Am Vet Med Assoc 1991;198:241-244.
- 3. Dubey JP, Carpenter JL, Speer CA, Topper MJ, Uggla A. Newly recognized fatal protozoan disease of dogs. J Am Vet Med Assoc 1988;192:1269-1285.
- 4. Dubey JP, Hattel L, Lindsay DS, Topper MJ. Neonatal Neospora caninum infection in dogs: isolation of the causative agent and experimental transmission. J Am Vet Med Assoc 1988;193:1259-1263.
- 5. Gondim LF, Pinheiro AM, Santos PO, Jesus EE, Ribeiro MB, Fernandes HS, Almeida MA, Freire SM, Meyer R, McAllister MM. Isolation of Neospora caninum from the brain of a naturally infected dog, and production of encysted bradyzoites in gerbils. Vet Parasitol 2001;101:1-7.
- 6. Gondim LF, McAllister MM, Anderson-Sprecher RC, Björkman C, Lock TF, Firkins LD, Gao L, Fischer WR. Transplacental transmission and abortion in cows administered Neospora caninum oocysts. J Parasitol 2004;90:1394-1400.
- 7. Pinheiro AM, Costa SL, Freire SM, Meyer R, Almeida MA, Tardy M, El-Bachá R, Costa MF. Astroglial cells in primary culture: a valid model to study Neospora caninum infection in the CNS. Vet Immunol Immunopathol 2006;113:243-247.
- 8. Pinheiro AM, Costa SL, Freire SM, Meyer R, Almeida MA, Tardy M, El-Bachá R, Costa MF. Neospora caninum: infection induced IL-10 overexpression in rat astrocytes in vitro. Exp Parasitol 2006;112:193-197.
- 9. Pinheiro AM, Costa SL, Freire SM, Ribeiro CS, Tardy M, El-Bachá RS, Costa MF. Neospora caninum: early immune response of rat mixed glial cultures after tachyzoites infection. Exp Parasitol 2010;124:442-447.
- 10. Manach C, Scalbert A, Morand C, Rémésy C, Jiménez L. Polyphenols: food sources and bioavailability. Am J Clin Nutr 2004;79:727-747.
- 11. De Clercq E. Current lead natural products for the chemotherapy of human immunodeficiency virus (HIV) infection. Med Res Rev 2000;20:323-349.
- 12. van der Watt E, Pretorius JC. Purification and identification of active antibacterial components in Carpobrotus edulis L. J Ethnopharmacol 2001;76:87-91.
- 13. Youn HJ, Lakritz J, Kim DY, Rottinghaus GE, Marsh AE. Anti-protozoal of medicinal herb extracts against Toxoplasma gondii and Neospora caninum. Vet Parasitol 2003;116:7-14.
- 14. Jean-Moreno V, Rojas R, Goyeneche DH, Coombs GH, Walker J. Leishmania donovani: differential activities of classical topoisomerase inhibitors and antileishmanials against parasite and host cells at the level of DNA topoisomerase 1 and in cytotoxicity assays. Exp Parasitol 2006;112:21-30.
- 15. Cortázar TM, Coombs GH, Walker J. Leishmania panamensis: comparative inhibition of nuclear DNA topoisomerase II enzymes from promastigotes and human macrophages reveals anti-parasite selectivity of fluoroquinolones, flavonoids and pentamidine. Exp Parasitol 2007;116:475-482.
- 16. Fonseca-Silva F, Inacio JD, Canto-Cavalheiro MM, Almeida-Amaral EE. Reactive oxygen species production and mitochondrial dysfunction contribute to quercetin induced death in Leishmania amazonensis. PLoS One 2011;6:e14666.
- 17. Arioka S, Sakagami M, Uematsu R, Yamaguchi H, Togame H, Takemoto H, Hinou H, Nishimura S. Potent inhibitor scaffold against Trypanosoma cruzi trans-sialidase. Bioorg Med Chem 2010;18:1633-1640.
- 18. Assis DM, Gontijo VS, de Oliveira Pereira I, Santos JA, Camps I, Nagem TJ, Ellena J, Izidoro MA, dos Santos Tersariol IL, de Barros NM, Doriguetto AC, dos Santos MH, Juliano MA. Inhibition of cysteine proteases by a natural biflavone: behavioral evaluation of fukugetin as papain and cruzain inhibitor. J Enzyme Inhib Med Chem 2013;28:661-670.
- 19. Caldas LA, Seabra SH, Attias M, de Souza W. The effect of kinase, actin, myosin and dynamin inhibitors on host cell egress by Toxoplasma gondii. Parasitol Int 2013;62:475-482.
- 20. Nones J, Stipursky J, Costa SL, Gomes FC. Flavonoids and astrocytes crosstalking: implications for brain development and pathology. Neurochem Res 2010;35:955-966.
- 21. Silva AR, Pinheiro AM, Souza CS, Freitas SR, Vasconcellos V, Freire SM, Velozo ES, Tardy M, El-Bachá RS, Costa MF, Costa SL. The flavonoid rutin induces astrocyte and microglia activation and regulates TNF-alpha and NO release in primary glial cell cultures. Cell Biol Toxicol 2008;24:75-86.
- 22. Cookson MR, McClean R, Pentreath VW. Preparation and use of cultured astrocytes for assay of gliotoxicity. Methods Mol Biol 1995;43:17-23.
- 23. Mosmann T. Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays. J Immunol Methods 1983;65:55-63.
- 24. Rosenfeld G. Corante pancrômico para hematologia e citologia clínica: nova combinação dos componentes do may-grunwald e do giemsa num só corante de emprego prático. Mem Inst Butantan 1947;20:329-335.
- 25. Wang Z, Li DD, Liang YY, Wang DS, Cai NS. Activation of astrocytes by advanced glycation end products: cytokines induction and nitric oxide release. Acta Pharmacol Sin 2002;23:974-980.
- 26. Jesus EE, Pinheiro AM, Santos AB, Freire SM, Tardy MB, El-Bachá RS, Costa SL, Costa MF. Effects of IFN-γ, TNF-α, IL-10 and TGF-β on Neospora caninum infection in rat glial cells. Exp Parasitol 2013;133:269-274.
- 27. Manning P, Cookson MR, McNeil CJ, Figlewicz D, Shaw PJ. Superoxide-induced nitric oxide release from cultured glial cells. Brain Res 2001;911:203-210.
Fig. 1Effect of flavonoids and Neospora caninum infection on the metabolism of glial cell cultures measured using the MTT assay. (A, B) MTT test absorbance (ABS) at 540 nm in cultured cells exposed to flavonoids (50 µM) for (A) 24 hr and (B) 48 hr of no infection with NC-1. (C) MTT test absorbance at 540 nm in cells exposed to flavonoids (50 µM) for 48 hr that were infected with NC-1 (+NC-1) during the last 24 hr of treatment. The absorbance results are expressed as the mean percentage±SD from the control (0.1% DMSO), which was considered 100%. *Significance was based on Dunnett's Multiple Comparison test.

Fig. 2Effect of flavonoids on Neospora caninum infection in glial cell cultures. (A) NC-1 tachyzoites in the cell lysates were quantified 24 hr after infection under control conditions (0.1% DMSO) and in flavonoid-pretreated cultures. Control cultures were considered 100%. (B) The proportion of vacuoles determined in relation to the total number of cells analyzed per field. NC-1 parasitophorous vacuoles were identified by immunocytochemistry followed by Rosenfeld staining in cultures under control conditions (0.1% DMSO) and in the cells pretreated with flavonoids. (C) Photomicrograph of NC-1 parasitophorous vacuoles (arrows) in control cultures. Insert showing NC-1 parasitophorous vacuoles at a larger scale (scale bar=10 µm). (D) Photomicrograph of NC-1 parasitophorous vacuoles (arrow) in cultures pretreated with flavonoid F1.

Fig. 3Effect of flavonoid treatment and Neospora caninum infection in glial cell cultures on production of nitric oxide (NO). Determination of NaNO2 in the culture medium of cells under control conditions (0.1% DMSO) or treated with flavonoids (50 µM) with or without NC-1 infection. *Statistical significance based on the Dunnett's Multiple Comparison test.