Abstract
Paleopathologists have begun exploring the pathoecology of parasitic diseases in relation to diet and environment. We are summarizing the parasitological findings from a mummy in the site of Lapa do Boquete, a Brazilian cave in the state of Minas Gerais. These findings in context of the archaeology of the site provided insights into the pathoecology of disease transmission in cave and rockshelter environments. We are presenting a description of the site followed by the evidence of hookworm, intestinal fluke, and Trypanosoma infection with resulting Chagas disease in the mummy discovered in the cave. These findings are used to reconstruct the transmission ecology of the site.
-
Key words: Chagas disease, hookworm, Echinostoma, cave, prehistory, Brazil
INTRODUCTION
Pathoecology is the study of past behavioral and environmental determinants of infection [
1–
6]. Examples of behaviors include crowding, sanitation, hygiene, and trade. Examples of environmental determinants include presence of pathogens, infection reservoirs, intermediate hosts as well as climate. These features of prehistoric life were affected by environmental factors, such as climate and soil conditions. Pathoecology began to emerge in the Southwest USA with the establishment of a link between the emergence of parasitic infections and Ancestral Pueblo cultural development [
7]. This pathoecology approach was based on coprolite and mummy studies. Such studies resulted in the recovery of specific infection organisms, each with its own life cycle requirements. These life cycle requirements shed light on the specific aspects of behavior and environment that existed at sites in prehistory.
Analyses of Andean mummies and coprolites contributed to the emergence of pathoecology as a concept and applied on a population scale. Martinson and her colleagues [
1] codified the concept of ‘pathoecology’ to explain patterns of parasitic infections in archaeological sites in the Moquegua Valley of southern Peru. They analyzed mummies and coprolites and developed pathology profiles of 4 contemporaneous villages associated with the same archaeological culture [
1,
8]. This research showed that the parasitism at several villages was defined by occupation, trade, status, presence of domestic animals, and site location relative to fresh water access. At the same sites, Reinhard and Buikstra [
8] analyzed the epidemiological diversity of head lice infection and were able to relate high infestations to specific male-associated activities, such as elaborate hair styles and use of headwear. Additional research in Chile showed that aggregation imposed by the Inca on local populations elevated crowd infections [
9]. These Andean studies were used to develop a pathoecological study from analysis of many mummies and coprolites excavated from several archaeological sites. We are presenting below an example of how pathoecology can be reconstructed from a single mummy exhibiting multiple infections.
Reinhard and Bryant [
6] and Reinhard [
2,
3] developed the theory of pathoecology to include Pavlovsky’s [
10] concept of nidality in identifying foci of prehistoric infection. He combined ecological factors into a predictive tool for infection. He included vectors, reservoir hosts, humans, and favorable external environments. He defined a nidus as that portion of a natural geographic landscape which contains a community consisting of a pathogen, vectors, reservoir hosts, and recipient hosts, and possessing an environment in which the pathogen can circulate. He further found that pathogens possessed nidality. Nidality is the characteristic of an infectious agent to occur in distinct nidi, such as being associated with particular geographic, climatic or ecological conditions. Reinhard [
2] applied Pavlovsky’s concept to tracing interior and exterior nidi at Chacoan Greathouses in New Mexico. Reinhard and Bryant [
6] and Reinhard [
3] incorporate environmental and dietary data to explain how environmental collapse exacerbates the nidality of infectious diseases. White and her colleagues [
11] applied pathoecology on a regional scale to explain patterns of scurvy and anemia in Belize using stable isotopic data combined in an environmental perspective. Most recently, Reinhard and Araújo [
5] applied pathoecology to devised excavation strategies to test hypotheses regarding
Trypanosoma cruzi transmission in the Lower Pecos Canyonlands of Texas and Coahuila. In that region,
T. cruzi infection exists in a sylvatic cycle involving woodrats (
Neotoma spp.) and triatomine insects (
Triatoma spp). The area’s prehistoric hunter-gatherer subsistence strategy impacted this life cycle directly through human predation on wood rats and through the construction of baking pits which amplified woodrat/triatomine habitat. That expanded the habitat suitable for both woodrats and triatomine insects. The construction of baking features in rock shelters used by humans resulted in the close association of triatomines with humans [
14]. They proposed field excavation that would test the nidi of transmission for triatomine insects and woodrat bone. Via molecular testing of recovered bones and insects, the transmission of
T. cruzi would be explored.
Recently, pathoecology has benefited from new diagnostic methods that expand our ability to recover evidence of parasites. This is illustrated by the combination of molecular biology, microscopy, and radiographic diagnosis of gross pathology applied to a mummy from Minas Gerais, Brazil.
ARCHAEOLOGICAL BACKGROUND
The northern region of Minas Gerais is a transitional zone between
cerrado (savanna) and
caatinga (desert vegetation). Forest formation is present in some areas, specifically along water courses [
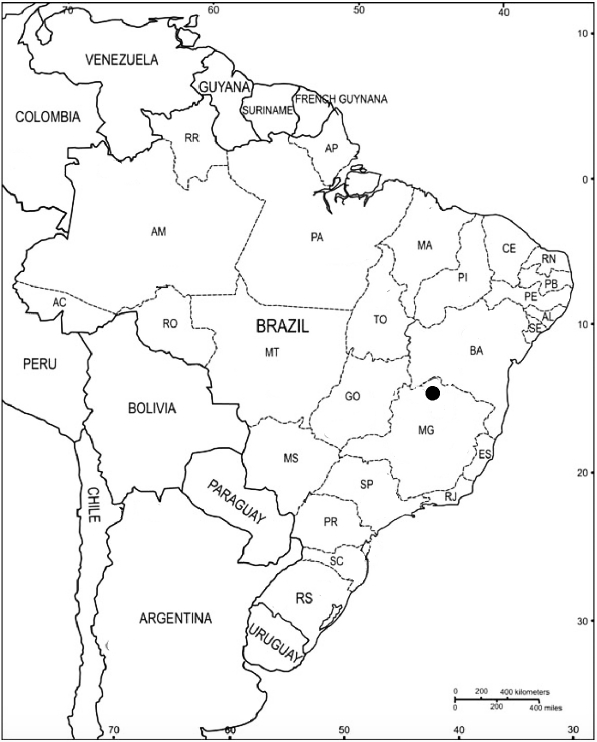
12]. The Site of Lapa do Boquete is located in a karstic canyon, the Peruaçu Valley, in the northern region of Minas Gerais state, Brazil (
Fig. 1). The Peruaçu River’s origin is located on gneiss bedrock, on the bank of the Rio São Francisco. Its middle course cuts through Precambrian calcareous formations, and in the past it was almost entirely subterranean. The roof collapse exposed the river bed, forming a canyon with cliffs 50 to 100 m high, characterized by karstic forms, including sinkholes (dolines), and subterranean river sections extending 1 to 3 km in length [
13–
15].
Excavations at Lapa do Boquete followed natural and cultural stratigraphy. Sedimentological features like color, texture, and composition where used to define geologic strata, and each stratum was divided into levels according to cultural stratigraphy.
Décapage excavation was done, with 3-dimensional plotting of artifacts, features, and excavation units [
12,
15].
Nine strata were found [
16]. Stratum 0 dated to the historic period and was composed of 5 to 15 cm of sediments mixed with modern dry cattle manure, and great quantities of archaeological material. Stratum 1 dates between 500 and 2,000 years ago. It was characterized by many hearths and abundant lithic artifacts. Strata 2 and 3 date between 2,000 and 6,000 years ago [
16]. Both of these strata were heavily disturbed by the activity of prehistoric groups that occupied the cave during later periods, and were disturbed by intrusive circular pits of 20 to 40 cm in depth. These pits were dug in order to bury baskets containing vegetal material, such as maize, fruits, and nuts. The base of Stratum 3 has been dated to 5,960±100 B.P. The remaining strata date to earlier Holocene occupations between 6,000 and 12,000 years ago and are characterized by lithic and faunal materials [
16]. These earlier strata are not immediately relevant to the discovery of parasite infections and will not be discussed further.
Kipnis [
16] described the archaeological remains. Ceramic fragments were recovered from Strata 0 and 1. The ceramics belong to the
Una tradition, which is characterized by thin-walled, undecorated small vessels found throughout Central Brazil, and almost exclusively in caves or rockshelters. Some of the other material culture found in the upper levels are a complete axe handle made of wood, beads made out of bone and wood, spatula-like tools made out of deer metapodials, and large planes used for woodworking made out of land snail (Strophocheilidae) shells. The latter 2 are relatively common in all levels. Stratum 1 contained 28 plant taxa [
16]. These included cultivated plants, such as
Gossypium sp. (cotton),
Manihot sp. (manioc),
Phaseolus (bean), and
Zea mays (maize).
Six human burials were found at Lapa do Boquete. One in stratum 1, another in stratum 2, and 4 in the older strata. Chronologically, 3 of the 6 burials range from ca. 7,000 to ca. 4,500 years ago, and the other 3 range from ca. 1,200 to ca. 600 years ago [
17]. One male adult and a child (10–14 years old) were buried between 600 and 1,000 years ago. The adult male was dated by radiocarbon method to 560±40 years ago [
18]. This is the focus of the studies in this review.
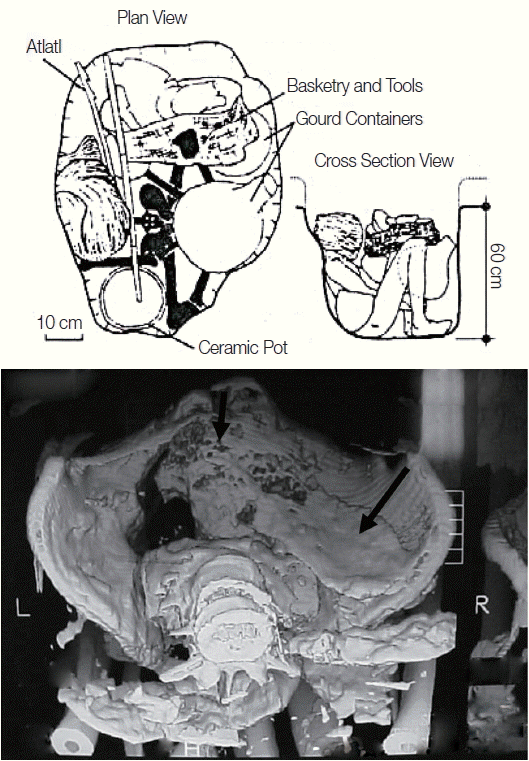
The body of the adult male was loosely flexed, wrapped in plant leaves, and buried with baskets (
Fig. 2). The man’s head was wrapped in plant fibers. Numerous artifacts were buried with him, including a dart thrower (atlatl), containers made of gourds, a ceramic pot, and numerous tools [
17]. The arms, legs, abdominal skin, and some muscles were well preserved. Although healed fractures of the foot phalanges were identified, there were no skeletal signs of violence. Lumbar spondylosis was observed. Researchers observed pronounced dental wear, caries, and dental abscesses. A large fecal mass, illustrated via ct-scan [
18,
19] (
Fig. 2), suggested that the individual experienced megacolon at the time of death.
PARASITOLOGICAL DIAGNOSES
Ferndandes and his colleagues [
18] analyzed 2 samples of bone (rib and metacarpus) and 5 samples of soft tissue from different places of the body for
T. cruzi DNA. The sample included tissue in contact with a coprolite. Researchers successfully recovered
T. cruzi genotype I from the individual [
18,
20]. This study linked the gross pathology of Chagas disease directly with the infectious agent. This landmark study is the first to link pre-Columbian
T. cruzi with Chagas disease in Brazil. It is likely that the individual had a broadly dispersed parasitic load because
T. cruzi I DNA was found in all tissue samples.
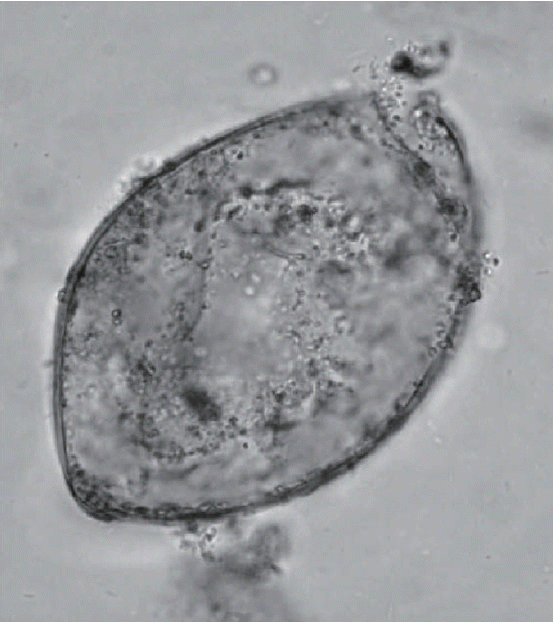
Another ground-breaking aspect of the Peruaçu study was the molecular refinement of microscopic diagnosis of trematode eggs (
Fig. 3). Coprolites were present in the mummy and were analyzed by Sianto and her colleagues [
19]. Eggs of a common intestinal parasite, hookworm, and an unusual intestinal parasite,
Echinostoma spp. were found. Hookworm eggs were relatively rare in this sample, and only 5 were observed.
Echinostoma spp. eggs were abundant and numbered 8,300 eggs per gram of coprolite. This suggested that the individual had a true infection. The morphology of the eggs could be described with certainty and were consistent the
Echinostoma genus. The species level diagnosis could not be reached with certainty, but the eggs were most consistent with those of
Echinostoma luisreyi. Leles and her colleagues [
21] showed that the DNA sequence of the ancient eggs was consistent with the species
Echinostoma paraensei. Therefore, molecular biology led to a more refined diagnosis at the species level.
PATHOECOLOGY IN THE PERUAÇU VALLEY
Echinostoma includes common intestinal flukes of aquatic birds and mammals. In this case, the species of
Echinostoma that infected the mummified man is most likely
E. paraensei [
19,
21]. The life cycle for the genus is complex involving snails as the first intermediate hosts. The second intermediate host is unknown, but could be other snails, leeches, fish, or tadpoles. In this intermediate host, they remain encysted until the intermediate host is eaten by a definitive host. In the case of
E. paraensei, a definitive host is
Nectomys squamipes, the South American water rat. This is an omnivorous mammal with a diet ranging from fruits and leaves to insects and small fish.
Various vertebrates and snails can be found throughout the year in the Peruaçu Valley region that serve as intermediate and definitive Echinostoma host. Human predation of snails and fish are the likely cause of the infection evidenced by the mummy. The infection nidi were water sources where human activity overlapped with the intermediate hosts that were eaten by humans.
Hookworm infection has a great antiquity in Brazil, among hunter-gatherers and horticulturalists [
21]. Reinhard [
2] presented a case that irrigation promoted hookworm infection among people who practiced agriculture. The best soils for survival are sandy or loamy. They require shady and wet conditions because they are killed by desiccation and heat. This is especially true for the hookworm species known from the prehistoric New World,
Ancylostoma duodenale. The nidi for infection were areas contaminated with human feces. This could have included areas immediately surrounding, or even within, human habitations.
T. cruzi has a moderately complex life cycle [
5,
18,
20,
23,
24]. The intermediate hosts are insects of the subfamily Triatominae. These insects feed on blood by biting vertebrates. Intracellular amastigotes destroy the intramural neurons of the autonomic nervous system in the intestine and heart. The megacolon evident on the Lapa do Boquete mummy was caused by this destruction of the large intestine neurons. The peristalsis is disrupted, and the colon fills with feces.
T. cruzi also exhibits transplacental or transmammary infection. Infection can also occur by eating food contaminated with trypomastigotes, either in the form of triatomine feces in plant foods or blood from incompletely cooked meat.
For ancient
T. cruzi infection in Texas cave settings, Reinhard and his colleagues [
25] hypothesized that several pathoecological aspects of human, parasite, and vector interaction caused the infection in prehistoric peoples. Prehistoric people frequently deposited large amounts of vegetal material in their rockshelter habitations as discovered in Lapa do Boquete. This created a good environment for triatomine bugs which transmit
T. cruzi. Humans worked and slept in the natural shelter provided by the cave. Thus, they were in close proximity to the bugs which promoted infection. Eating raw meat from infected reservoir hosts, including rodents and armadillos can cause infection [
26]. Analysis by Kipnis [
16] showed that armadillo meat was commonly eaten in a poorly cooked state. Armadillos are a reservoir host for
T. cruzi and the habit of eating armadillo meat may have caused some infections.
The pathoecology of the Lapa do Boquete mummy can be reconstructed from his parasites. The environment was moist and warm. The soils where the man walked and worked included shaded, warm, sandy-loamy deposits where hookworm larvae could remains active. The environment supported wetland adapted intermediate and definitive hosts for
Echinostoma spp. Snail shells of the family Strophocheilidae were commonly found in the site. However, species in this family are not recorded as
E. paraensei intermediate hosts. The snails
Biomphalaria glabrata and
Physa rivalis are
E. paraensei intermediate hosts [
27–
29]. Consumption of intermediate hosts, probably fish, was the source of
Echinostoma infection [
19]. There are no recorded cases of
E. paraensei infection in humans, so this is a provocative finding.
Each of these infections has a distinct nidality. T. cruzi has the most restricted nidi. A nidus for T. cruzi is an enclosed area where the infected triatomine is associated with sleeping humans. The caves in the Peruaçu Valley, including Lapa do Boquete, were the primary nidi for infection. In the case of infection by eating contaminated meat, the nidus extended over the area overlapped by human hunters and armadillos.
The nidus for hookworm infection must include soil somewhat close to fecal deposits where humans and infective larvae are active. Areas near latrines or agricultural fields contaminated with feces are common nidi for hookworm infection. The most common method to control for hookworms is to use footwear, but Behnke and colleagues [
30] document problems with the wearing of sandals among modern agriculturalists in Mali. They state [
30, p 352], “soil sticks to sandals, making them uncomfortable and frustrating to wear when tilling soil, and risking damage. As a result of this practice, those who often wore shoes still became infected through bare skin.” Lapa do Boquete inhabitants wore sandals or were barefoot. Thus, simple footwear left them open to infection. Relevant to Lapa do Boquete pathoecology, Schad and his colleagues [
31] documented another scenario for hookworm infection. They found that in West Bengal, villagers defecated in areas around the village peripheries. People used the same areas day after day for this purpose. As a result, hookworm larvae proliferated in those areas and the brief time spent by humans in the contaminated areas was sufficient for hookworm infection.
In conclusion, the spectrum of parasites from a single individual reveals a relatively complex pathoecology for his community of horticulturalists. Risk of hookworm infection was associated with fecal contamination in moist areas without adequate foot protection. Echinostome infection resulted from the consumption of intermediate host fish or mollusks. T. cruzi infection may have cycled in sleeping areas or through ingestion of reservoir hosts.
Notes
-
CONFLICT OF INTEREST
We declare no conflict of interest related to this study.
ACKNOWLEDGMENTS
The writing of this manuscript was supported by the Brazilian National Council for Scientific and Technological Development (CNPq), Brazilian Improving Coordination of Higher Education Personnel (CAPES), and the Fulbright Commission.
Fig. 1Map of Brazil showing the location of the Lapa do Boquete and the Peruaçu Valley.
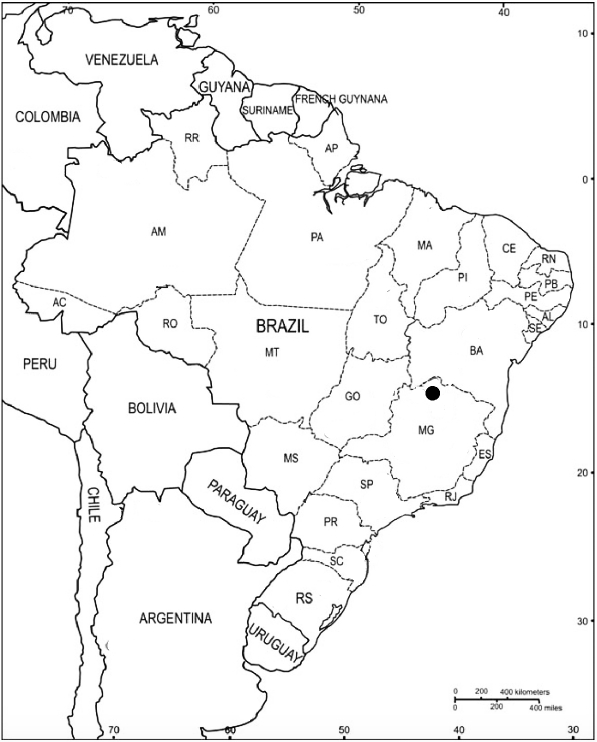
Fig. 2The upper image is redrafted from Prous and Schlobach [
17, pp 18–19]. The plan view shows the individual’s bones in solid black. The head was wrapped in leaf foliage. A dart thrower (seteira in Portuguese, atlatl in archaeological parlance), basketry, gourd containers, and a ceramic vessel were buried this the individual. The cross section of the burial shows the man’s placement relative to major artifacts. The lower image shows the CT-scan of the pelvic girdle showing a large fecal mass indicated by the arrow.

Fig. 3Echinostome egg as illustrated in Sianto’s original article describing the egg morphology [
19].

References
- 1. Martinson E, Reinhard KJ, Buikstra JE, de la Cruz KD. Pathoecology of Chiribaya parasitism. Mem Inst Oswaldo Cruz 2003;98( suppl 1):195-205.
- 2. Reinhard K. Parasite pathoecology of Chacoan Great Houses: the healthiest and wormiest Ancestral Puebloans. In: Reed PF, editor. Chaco’s Northern Prodigies Salmon, Aztec, and the Ascendancy of the Middle San Juan Region after AD 1100. Salt Lake City, Utah, USA. University of Utah Press; 2008. p. 86-95.
- 3. Reinhard KJ. Pathoecology of two Ancestral Pueblo villages. In: Reitz EJ, Scarry CM, Scudder SJ, editors. Case Studies in Environmental Archaeology. 2nd ed. New York, USA. Plenum Press; 2008. p. 191-210.
- 4. Reinhard KJ, Araújo A. Prehistoric earth oven facilities and the pathoecology of Chagas disease in the Lower Pecos Canyonlands. J Archaeol Sci 2015;53:227-234.
- 5. Reinhard KJ, Araújo A. Synthesizing archaeology with parasitology in paleopathology. In: Buikstra J, Roberts C, editors. A Global History of Paleopathology. Oxford, UK. Oxford University Press; 2012. p. 751-764.
- 6. Reinhard KJ, Bryant VM. Pathoecology and the future of coprolite studies. In: Stodder AWM, editor. Reanalysis and Reinterpretation in Southwestern Bioarchaeology. Tempe, Arizona, USA. Arizona State University Press; 2008. p. 199-216.
- 7. Reinhard KJ. The cultural ecology of prehistoric parasitism on the Colorado Plateau as evidenced by coprology. Am J Phys Anthropol 1988;77:355-366.
- 8. Reinhard KJ, Buikstra JE. Louse infestation of the Chiribaya Culture, Southern Peru: variation in prevalence by age and sex. Mem Inst Oswaldo Cruz 2003;98:173-179.
- 9. Santoro C, Vinton SD, Reinhard KJ. Inca expansion and parasitism in the Lluta Valley: preliminary data. Mem Inst Oswaldo Cruz 2003;98:161-163.
- 10. Pavlovsky EN. Natural Nidality of Transmissible Diseases with Special Reference to the Landscape Ecology of Zooanthroponoses. Plous FK Jr, translator. Urbana, Illinois, USA. University of Illinois Press; 1966. p 248.
- 11. White C, Maxwell J, Dolphin A, Williams J, Longstaffe F. Pathoecology and paleodiet in postclassic/historic Maya from northern coastal Belize. Mem Inst Oswaldo Cruz 2006;101( suppl 2):35-42.
- 12. Moura MT. A evolução do sítio arqueológico Lapa do Boquete na paisagem cárstica do vale do rio Peruaçu: Januária (MG). Masters Thesis. Departamento de Geografia, Universidade de São Paulo; 1997. 208.
- 13. Piló LB. Rochas carbonáticas e relevos cársticos em Minas Gerais. O Carste 1997;9:72-78.
- 14. Araújo AG, Piló LB, Neves WA, Atui JP. Human occupation and paleoenvironments in South America: expanding the notion of an “Archaic Gap”. Rev Mus Arqueol Etnol São Paulo 2015;15&16:3-35.
- 15. Prous A. Aspectos arqueológicos do vale do Peruaçu. Congresso da Associacção Brasileira de Estudos do Quaternário, 3. Belo Horizonte, Brazil: Roteiro de Excursão. Imprensa Universitária da UFMG; 1991. 66-70.
- 16. Kipnis R. Foraging societies of Eastern Central Brazil: an evolutionary ecological study of subsistence strategies during the Terminal Pleistocene and Early/Middle Holocene. PhD dissertation. Michigan, USA. University of Michigan; 2002. p. 1-620.
- 17. Schlobach M, Prous A. Sepultamentos pré-históricos do Vale do Peruaçu. Rev Mus Arqueol Etnol São Paulo 1997;7:3-21.
- 18. Fernandes A, Iñiguez AM, Lima VS, Souza SM, Ferreira LF, Vicente AC, Jansen AM. Pre-Columbian Chagas disease in Brazil: Trypanosoma cruzi I in the archaeological remains of a human in Peruaçu Valley, Minas Gerais, Brazil. Mem Inst Oswaldo Cruz 2008;103:514-516.
- 19. Sianto L, Reinhard KJ, Chame M, Chaves S, Mendonça S, Gonçalves ML, Fernandes A, Ferreira LF, Araújo A. The finding of Echinostoma (Trematoda: Digenea) and hookworm eggs in coprolites collected from a Brazilian mummified body dated of 600–1,200 years before present. J Parasitol 2005;91:972-975.
- 20. Macedo AM, Machado CR, Oliveira RP, Pena SD. Trypanosoma cruzi: genetic structure of populations and relevance of genetic variability to the pathogenesis of Chagas disease. Mem Inst Oswaldo Cruz 2004;99:1-12.
- 21. Leles D, Cascardo P, Freire Ados S, Maldonado A Jr, Sianto L, Araújo A. Insights about echinostomiasis by paleomolecular diagnosis. Parasitol Int 2014;63:646-649.
- 22. Reinhard K, Araújo A, Ferreira LF, Coimbra CE. American hookworm antiquity. Med Anthropol 2001;20:96-101.
- 23. Aufderheide AC, Salo W, Madden M, Streitz J, Buikstra J, Guhl F, Arriaza B, Renier C, Wittmers LE Jr, Fornaciari G, Allison M. A 9,000-year record of Chagas’ disease. Proc Natl Acad Sci USA 2004;101:2034-2039.
- 24. Lima VS, Iñiguez AM, Otsuki K, Ferreira LF, Araújo A, Vicente ACP, Jansen AM. Chagas disease by Trypanosoma cruzi lineage I in hunter-gatherer ancient population in Brazil. Emerg Infect Dis 2008;14:1001-1002.
- 25. Reinhard K, Fink TM, Skiles J. A case of megacolon in Rio Grande Valley as a possible case of Chagas Disease. Mem Inst Oswaldo Cruz 2003;98( suppl 1):165-172.
- 26. Dias JCP. Epidemiology of Chagas disease. In: Wendel S, Brener Z, Camargo MF, Rassi A, editors. Chagas Disease (American Trypanosomiasis): Its Impact on Transfusion and Clinical Medicine. São Paulo, Brasil. Sociedade Brasileira de Hematologia e Hemoterapia; 1992. p. 49-80.
- 27. Lie KJ, Basch PF. The life history of Echinostoma paraensei sp. n. (Trematoda: Echinostomatidae). J Parasitol 1967;53:1192-1199.
- 28. Loker ES, Bayne CJ, Yui MA. Echinostoma paraensei: hemocytes of Biomphalaria glabrata as targets of echinostome mediated interference with host snail resistance to Schistosoma mansoni. Exp Parasitol 1986;62:149-154.
- 29. Maldonado A Jr, Zeitone BK, Amado LA, Rosa IF, Machado-Silva JR, Lanfredi RM. Biological variation between two Brazilian geographical isolates of Echinostoma paraensei. J Helminthol 2005;79:345-351.
- 30. Behnke JM, De Clercq D, Sacko M, Gilbert FS, Ouattara DB, Vercruysse J. The epidemiology of human hookworm infections in the Southern Region of Mali. Trop Med Int Health 2000;5:343-354.
- 31. Schad GA, Nawalinski TA, Kochar V. Human ecology and the distribution and abundance of hookworm populations. In: Croll N, Cross L, editors. Human Ecology and Infectious Diseases. London, UK. Academic Press; 1983. p. 187-273.