Abstract
Giardia lamblia, an anaerobic, amitochondriate protozoan parasite causes parasitic infection giardiasis in children and young adults. It produces pyruvate, a major metabolic product for its fermentative metabolism. The current study was undertaken to explore the effects of pyruvate as a physiological antioxidant during oxidative stress in Giardia by cysteine-ascorbate deprivation and further investigation upon the hypothesis that oxidative stress due to metabolism was the reason behind the cytotoxicity. We have estimated intracellular reactive oxygen species generation due to cysteine-ascorbate deprivation in Giardia. In the present study, we have examined the effects of extracellular addition of pyruvate, during oxidative stress generated from cysteine-ascorbate deprivation in culture media on DNA damage in Giardia. The intracellular pyruvate concentrations at several time points were measured in the trophozoites during stress. Trophozoites viability under cysteine-ascorbate deprived (CAD) medium in presence and absence of extracellular pyruvate has also been measured. The exogenous addition of a physiologically relevant concentration of pyruvate to trophozoites suspension was shown to attenuate the rate of ROS generation. We have demonstrated that Giardia protects itself from destructive consequences of ROS by maintaining the intracellular pyruvate concentration. Pyruvate recovers Giardia trophozoites from oxidative stress by decreasing the number of DNA breaks that might favor DNA repair.
-
Key words: Giardia lamblia, oxidative stress, cysteine-ascorbate, antioxidant, pyruvate, reactive oxygen species, cytotoxicity
INTRODUCTION
Giardia lamblia, an enteric parasite causing giardiasis, has a simple life cycle consisting of encystation and excystation [
1]. It is responsible for foodborne and waterborne diarrhea worldwide [
2]. It imparts to an approximately 280 million symptomatic infections in human yearly [
3], and it has been included as part of WHO Neglected Disease Initiative since 2004 [
4]. The symptoms of giardiasis appear after 6–15 days of infection, and it is characterized by diarrhea or greasy stool, fatigue, nausea, bloating, abdominal cramps, excessive gas, and headache [
5]. The impact of clinical complications is high in case of malnourished children and immunodeficient individuals. Although it cannot cross intestinal lining and does not secrete any toxin, recent data suggest that
Giardia increases intestinal permeability by induction of apoptosis of intestinal epithelial cells. There are many drugs available, but metronidazole is the most commonly used drug for treatment. It is incapable to tolerate elevated oxygen pressure and in the upper intestine where this organism generally inhabits, the oxygen (O
2) concentration there has been measured at 60 μM [
6]. In addition to this, the amitochondriate parasite lacks some of the conventional enzymes that detoxify reactive oxygen species (ROS), like superoxide dis-mutase (SOD), catalase, peroxidase, and glutathione reductase [
7]. Cysteine is not incorporated de novo and is not synthesized from cystine. It appears to be imported into the cell by passive diffusion, although active transport may account for some of the acquisition of cysteine [
8]. Trophozoites containing free thiol (−SH) groups on their surface was described previously by the toxicity of thiol-blocking agents that are unable to penetrate intact cells [
9]. It suggests that the toxicity is due to the reaction between the agents and surface proteins [
10]. The trophozoites of
Giardia are also protected by ascorbic acid under high partial pressure of oxygen [
11]. Previously it was reported that cysteine-ascorbate deprivation generates ROS in
Giardia that had a tendency towards apoptotic death [
12]. Some of the genes which are differentially regulated during stress generated by cysteine-ascorbate deprived (CAD) medium are involved in different pathways of energy metabolism in
Giardia [
13].
Pyruvate, produced by glycolysis, is a crucial intermediate of energy metabolism in this microbe and is the precursor of most metabolic end products which are dependent on the ambient values for oxygen tension [
14]. In
G. lamblia, the conversion of acetate from pyruvate, a very important metabolic sequence of energy metabolism occurs in the cytoplasm. The energy of thioester bond of acetyl-CoA is always conserved by substrate level phosphorylation. Thus, acetate formation plays a very significant role in
Giardia’s energy metabolism. It has been hypothesized that change in redox state of NAD(P)H pools affects the relative rates of end product formation [
15]. The detailed mechanism by which the parasite could detoxify excess ROS produced during an oxidative stress is still unknown. In this study, we have demonstrated the effects of pyruvate on oxidative stress regulation and redox homeostasis in
G. lamblia under CAD medium stress.
MATERIALS AND METHODS
Chemicals and solutions
All reagents were purchased from Sigma-Aldrich (St. Louis, Missouri, USA) unless otherwise specified. Pyruvate (sodium salt), menadione, acetate, malate, α-ketoglutarate, mannitol, oxaloacetate, and other solutions were prepared freshly at the time of the experiment. For experimentation, same set of cells were taken from individual culture.
Culture methods
G. lamblia Portland1 strain (ATCCR 30888TM) trophozoites were maintained in TYIS-33 medium, supplemented with penicillin (100 U/ml), streptomycin (100 mg/ml), and 10% adult bovine serum. All the experiments were performed on trophozoites that had been harvested during the mid-exponential phase of growth by centrifugation at 2,000 rpm for 10 min and resuspended in PBS buffer (PH-7.2) containing 150 mM NaCl, 5 mM K2HPO4, and 1.8 mM KH2PO4. All the solutions were prepared freshly on the day of experiment performed. Same set of trophozoites were taken for an individual experimentation.
Confocal microscopy
ROS in trophozoites were assessed by using dichlorodihydrofluoresceine diacetate (H
2DCFDA) fluorescent probe, according to Schuessel et al. [
16] with few modifications. CAD medium treated trophozoites (2×10
6 cells/ml) were previously incubated with different metabolites (pyruvate, acetate, ascorbate etc.) at a concentration of 2 mM. H
2DCFDA, a membrane permeable dye and therefore we have not used any mild detergent for permeabilization and the cells were fixed with 2% paraformaldehyde after washing with PBS thrice. Consequently, observations were made with a confocal microscope (LSM510 Meta, Carl Zeiss, Thornwood, New York, USA).
Giardia trophozoites (107 cells/ml) were pre-incubated in the presence and absence of different concentration of pyruvate (0–5 mM) under cysteine-ascorbate deprivation. After 8 hr, trophozoites were washed and resuspended in phosphate buffered saline. ROS Levels were determined by spectrofluorometer (QuantaMaster 30, Photon Technology International, Birmingham, New Jersey, USA) at 530 nm after excitation at 488 nm.
Viability assessment by flowcytometric analysis
Treated and untreated Giardia trophozoites were previously incubated with or without pyruvate. Measurement of PI fluorescence with a Becton-Dickinson FACSARIA-III flowcytometer (BD Biosciences, San Jose, California, USA) was performed. Menadione (2-methyl-1, 4-napthoquinone) induced ROS generation was also determined in trophozoites previously incubated with or without pyruvate for 30 min. After 8 hr incubation, trophozoites were washed with PBS and added H2DCFDA (1.5 μM). The H2DCFDA loaded trophozoites were incubated in the dark at 37°C for 15 min. Then, the trophozoites were washed thrice with PBS and resuspended in PBS for flowcytometric analysis. All the data were analyzed by WinMDI 2.9 software (Scripps Institute, La Jolla, California, USA).
Thioburbituric acid (TBA) assay
The TBA assay was conducted to measure the MDA concentration as described by Bar-Or et al. [
17] with a slight modification. Malondialdehyde (MDA) is produced by the process of lipid peroxidation. This is a later product in the sequence of lipid peroxidation reactions [
18,
19]. It produces a chromophore when reacts with TBA. The absorbance of the chromophore was measured at 535 nm. The MDA concentration is presented as nmoles of MDA produced/mg protein using a molar extinction coefficient of 1.56×10
5 M
−1 cm
−1.
Intracellular pyruvate concentration was determined under cysteine-ascorbate deprivation after different time interval by using the Pyruvate Assay Kit (ab65342). Trophozoites (107 cells/ml) were incubated under CAD medium for 0 to 8 hr. Trophozoites were harvested and homogenization procedure was performed. Then, 6% perchloric acid (PA) was added to inactivate the enzyme. Supernatant was collected and used for pyruvate assay according to the manufacturer’s protocol.
DNA fragmentation assay
Untreated and CAD medium treated (for 8 hr at 35.5°C trophozoites were taken. The assay was done according to the Nandi et al. [
20] with little modification. After phenol-chloroform treatment, salt precipitation and 70% ethanol wash, the pellet was air dried and resuspended in sterile triple distilled water and run in a 1.5% agarose gel stained with ethidium bromide.
Determination of cell viability was studied by the use of Annexin-V FITC assay kit (IM3546). As CAD medium have been reported to induce DNA damage [
12], we have estimated the effects of pyruvate during incubation under CAD medium on DNA damage in
G. lamblia. Trophozoites were incubated for 8 hr under CAD medium with different substrates at same concentration (2 mM) (previously incubated for 30 min). Trophozoites were harvested and washed with ice-cold PBS and collected. After that, the assay was carried out according to the manufacturer’s protocol and analyzed by flowcytometry.
Treated and untreated trophozoites were harvested as per experimental procedure and the total RNA was extracted by TRIZOL (Invitrogen) method and gene expression studies were performed according to the Raj et al. [
13]. To understand the metabolic gene regulation of pyruvate metabolism pathway we have performed gene expression of PFOR (F: 5-ATCCAACGCGACCCAGAAG-3, R: 5-GTTCACTGCTTACTCCGCC-3); MDH (F: 5-GGAGACATGCTGGGCTACGA-3, R: 5-CGGCAGGAACCTCAAGCATA-3); PDK (F: 5-TGGAACCAACGATCTTACACAG-3, R: 5-GGAGACATAGTTCAGGCCAATC-3); PK (F: 5-CAGACCAGAAAAAGCACATCAG-3, R: 5-GCGGTCCATTCTTGAAAACTAC-3); ACS (F: 5-ACTGAGATCCTGGGGTACAAG-3, R: 5-ACGATGGACTCAAGGTAAAGG-3); ARGD (F: 5-GGCGAAGGCAAATGTTGAGT-3, R: 5-CGGACGATCGTGTAACCATTTT-3).
All the experiments were executed at least thrice in triplicates and the results were expressed as mean±standard error of the mean (SEM). Statistical analysis was evaluated by t-test or 1-way ANOVA followed by Kruskal-Wallis test (wherever applicable), using Graph Pad Prism software, version 4 (Graph-Pad Software, San Diego, California, USA); P <0.05 was considered as statistically significant.
RESULTS
Exogenously added pyruvate at physiological concentration attenuates intracellular ROS production in the trophozoites under cysteine-ascorbate deprivation
The H2DCFDA, a non-fluorescent molecule, is able to enter the cells. After getting entry into the cytosol of the cell, esterase activity renders the indicator, non-permeable by forming fluorescent product dichlorofluoresceine and the fluorescence intensity of the dye is proportional to the rate of oxidation by ROS.
Pyruvate can enter through the biological membrane in to the trophozoites and raise the level of intracellular pyruvate concentration. The H
2DCFDA assay was performed to study the intracellular generation of ROS in
G. lamblia in the presence and absence of pyruvate. Observation of cellular fluorescence in the trophozoites was examined by confocal microscopy under CAD medium stress conditions. Our results confirmed that exogenously added physiological concentrations of pyruvate attenuate fluorescence, produced by ROS under cysteine-ascorbate deprivation (
Supplementary Fig. 1).
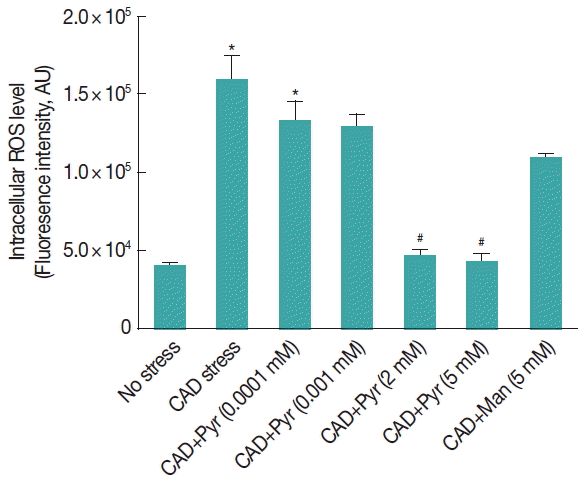
Trophozoites were previously incubated with increasing concentrations of pyruvate from 0.0001 to 5 mM and then exposed in CAD medium. The level of ROS was measured in
Giardia trophozoites with or without pyruvate. The ROS level increased significantly (
P <0.01) by cysteine-ascorbate deprivation than H
2DCFDA-loaded untreated trophozoites. The fluorescence intensity was lowered significantly (
P <0.05) in the presence of pyruvate for the range of concentrations from 2 to 5 mM (
Fig. 1).
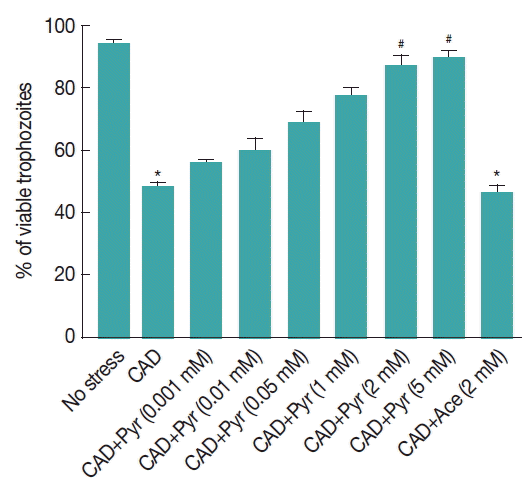
Pyruvate protects Giardia trophozoites from the toxicity induced CAD medium. Flowcytometric analysis was carried out to confirm the antioxidant activity of pyruvate in Giardia trophozoites. Trophozoites were incubated for 30 min with increasing concentration of sodium pyruvate under CAD medium for 8 hr.
Cysteine-ascorbate deprivation reduces trophozoites viability to 48.33%, which was significantly lower than the untreated trophozoites (93.6%,
P <0.001) (
Fig. 2). The trophozoites previously incubated with pyruvate (2 mM) and (5 mM) protects trophozoites significantly at 87.1% and 89.3% from CAD medium toxicity. When treated under CAD medium, trophozoites were protected by increasing concentrations of pyruvate.
Acetate, produced from pyruvate, did not vary the viability of Giardia either in control conditions or treated with CAD medium.
Pyruvate decreases intermediatory ROS generation by the induction of menadione in trophozoites during CAD medium stress
A progressive increase in the generation of ROS (as indicated by the enhancement in fluorescence intensity) with the incubation under CAD medium increased the rate of ROS generation. Prior incubation of H
2DCFDA loaded cells with increasing concentration of pyruvate (0.01 mM, 0.5 mM, and 2 mM) for 30 min attenuated the stressed induced fluorescence. Exposure of
Giardia trophozoites in increasing concentrations of menadione for 1 hr induced ROS generation that was significantly reduced by 2 mM sodium pyruvate (
Supplementary Fig. 2).
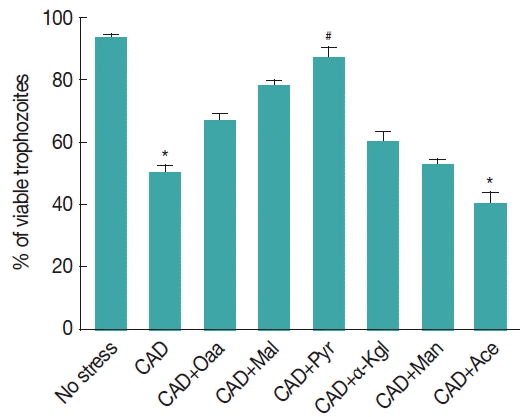
We have also observed that the preventive effects of pyruvate and other α-ketoacids against CAD medium stress. Trophozoites of Giardia were incubated under CAD medium for 8 hr after 30 min initial incubation in the presence of α-ketoglutarate and oxaloacetate, mannitol, malate, β-ketoglutarate. All the compounds were administered at a concentration of 2 mM.
In particular, acetate was observed to be ineffective, whereas oxaloacetate strongly prevented CAD medium cytotoxicity by 60.6%. Viability of trophozoites under CAD medium treated samples incubated with pyruvate and malate was restored to 88% (
P <0.01) and 78%, respectively; however, the viability of trophozoites were 53% and 40.6% when it was previously supplemented with mannitol and acetate, respectively, and was observed to be significantly lower than the control (
P <0.001) (
Fig. 3).
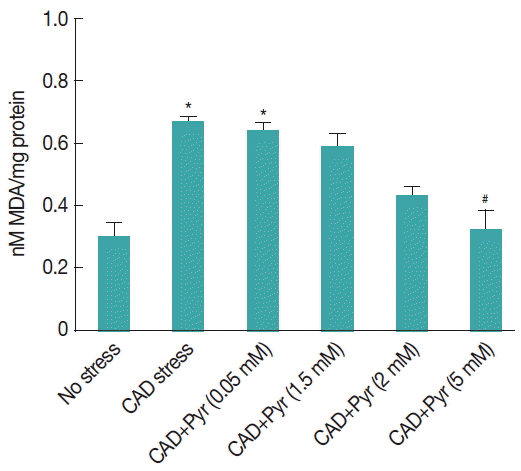
The degree of lipid peroxidation has been demonstrated on the basis of malondialdehyde (MDA) formation. We have measured lipid peroxidation status in G. lamblia under CAD medium stress with or without pyruvate supplementation. Lipid peroxidation was found to be increased significantly by 43% (P <0.01) in trophozoites under cysteine-ascorbate compared to the untreated trophozoites.
Supplementation of pyruvate (5 mM) significantly decreased the lipid peroxidation to 48% (
P <0.05) in the trophozoites under CA-deprived medium stress compared to the stressed trophozoites without pyruvate incubation (
Fig. 4).
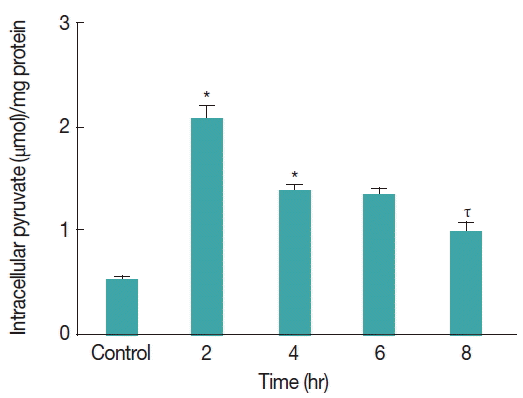
The intracellular pyruvate concentration in
G. lamblia trophozoites was measured during CAD medium induced stress conditions. Pyruvate content was quantified in trophozoites under CAD medium stress. Intracellular pyruvate concentration was observed to be increasing significantly from 0.5 μmol/mg proteins to 2.1 μmol/mg proteins (
P <0.001) after 2 hr under cysteine-ascorbate deprivation in
Giardia trophozoites (
Fig. 5). However, with time pyruvate concentration was decreased to 1.35 μmol/mg proteins from 2.1 μmol/mg proteins between 2nd and 4th hour and it remained almost same 1.3 μmol/mg proteins at 4th and 6th hour. Interestingly, after 8 hr of stress under cysteine-ascorbate deprivation, the concentration of pyruvate was significantly lowered to 0.97 μmol/mg proteins (
P <0.01). However, the trophozoites seemed to adjust between production and utilization of pyruvate in order to maintain the intracellular pyruvate level at and around 0.5 μmol/mg proteins (the normal pyruvate concentration in the trophozoites) during cysteine-ascorbate deprivation.
The hallmark of apoptosis in mammalian cell is the degradation of genomic DNA. Therefore, we investigated the DNA fragmentation pattern for untreated and stressed-induced trophozoites and also in cysteine-ascorbate stress-induced trophozoites supplemented with pyruvate. The stressed-induced trophozoites showed a DNA fragmentation pattern after 8 hr exposure to CAD medium. The ladder pattern was not clear as a metazoan DNA ladder and showed some degree of smearing with fragmented DNA in the low molecular weight region, identified by electrophoresis on a 1.5% agarose gel (
Supplementary Fig. 3).
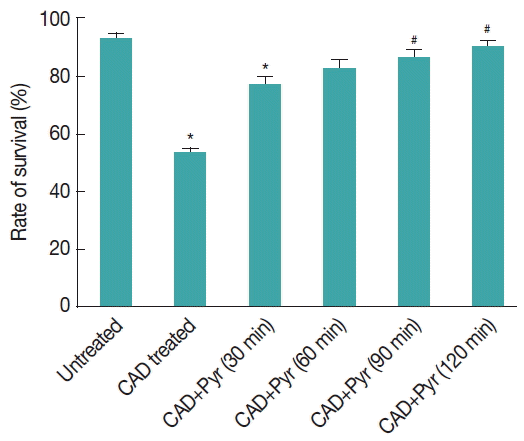
Externalization of phosphatidylserine upon exposure under CAD medium. We observed that the rate of recovery of the stressed
Giardia trophozoites increased with the increase in pyruvate concentration. In these recovery studies the major goal for administering pyruvate was to provide the substrate distal to glycolysis, thereby boosting ATP generation and acetate production. We tried to rescue trophozites of
G. lamblia by adding a pyruvate. Survival rate was estimated 24 hr later. We have determined that with time, the rate of recovery has increased for pyruvate after cysteine-ascorbate deprivation stress (
Fig. 6).
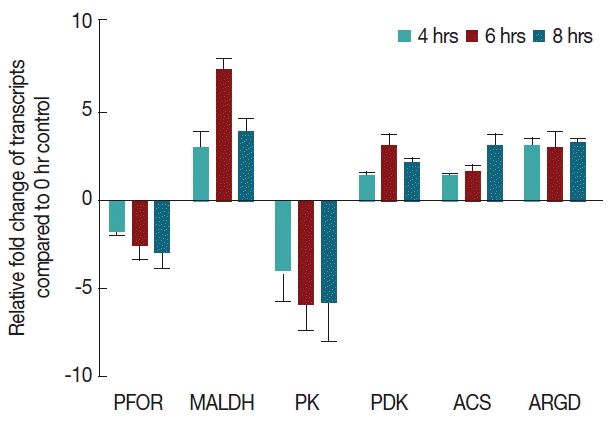
To understand the effect of cysteine-ascorbate deprivation stress in transcriptional regulation of gene expression in
G. lamblia, we performed a time course analysis of transcriptional regulation of pyruvate metabolism pathway under CAD medium stress using a quantitative RT-PCR. We have chosen 6 genes, related to the pyruvate metabolism of
G. lamblia modulated by at least 2 folds at one or more time points in response to cysteine-ascorbate deprivation (
Fig. 7).
In our study, we have shown that arginine deiminase (ARGD)-encoding gene was up-regulated in Giardia trophozoites during CAD medium stress. It suggests that Giardia needs energy to continue the process of stress management for its survival. In G. lamblia, pyruvate can be produced by 3 different pathways. Malate dehydrogenase (MDH) gene was up regulated at one or more time points upon cysteine-ascorbate deprivation. The gene showed a maximum induction at 6th hour of cysteine-ascorbate deprivation to maintain the pyruvate concentration at intracellular milieu. In response to cysteine-ascorbate deprivation pyruvate kinase remained down-regulated after 4th hour time points. The enzyme acetyl-CoA synthase transcript was up-regulated during CAD medium stress and increased the production of acetate, a lipid radical scavenger.
In our study, pyruvate ferredoxin oxidoreductase (PFOR)-encoding gene was down-regulated under CAD medium stress and reduced the conversion from acetyl-coA to acetate. However, cysteine-ascorbate deprivation has been observed to significantly harmonize the metabolic flux across the pyruvate metabolism in G. lamblia.
DISCUSSION
The present study reveals that pyruvate administration protects Giardia trophozoites against the cytotoxicity induced by CAD medium. It was previously stated that CAD medium produces ROS in Giardia. To establish its pathogenesis, Giardia has to build up an antioxidant protective mechanism.
The antioxidant protective effect of α-ketoacids has already been studied both in vitro in several cell types [
21] and in vivo in whole organs such as heart [
22]. It is well-known that pyruvate nonenzymatically reacts with hydrogen peroxide to produce acetate, CO
2, H
2O [
23]. With the help of the H
2DCFDA-based assays, intracellular production of ROS by
G. lamblia was monitored [
24].
CAD medium produces intracellular ROS in Giardia and the fluorescence was also increased by the addition of synthetic quinone menadione. Our results proved that intracellular generation of ROS was reduced by the supplementation of pyruvate and it also attenuated the menadione induced ROS generation. It was also shown to reduce the generation of total fluorescence arising from the oxidation of the H2DCFDA by spectrofluorometer and may also have acted a direct antioxidant role in Giardia.
The protective effect of pyruvate was reconstructed by several α-ketoacids, which share with pyruvate the ability to react with intracellular ROS; these compounds include mannitol, which is not an energy substrate. We have noticed that pyruvate has a higher capacity to scavenge ROS. It has been reported that elevated ROS production is very much related to a decline in the production of ATP as well as protein synthesis and increase in DNA breakage and lipid peroxidation [
25,
26].
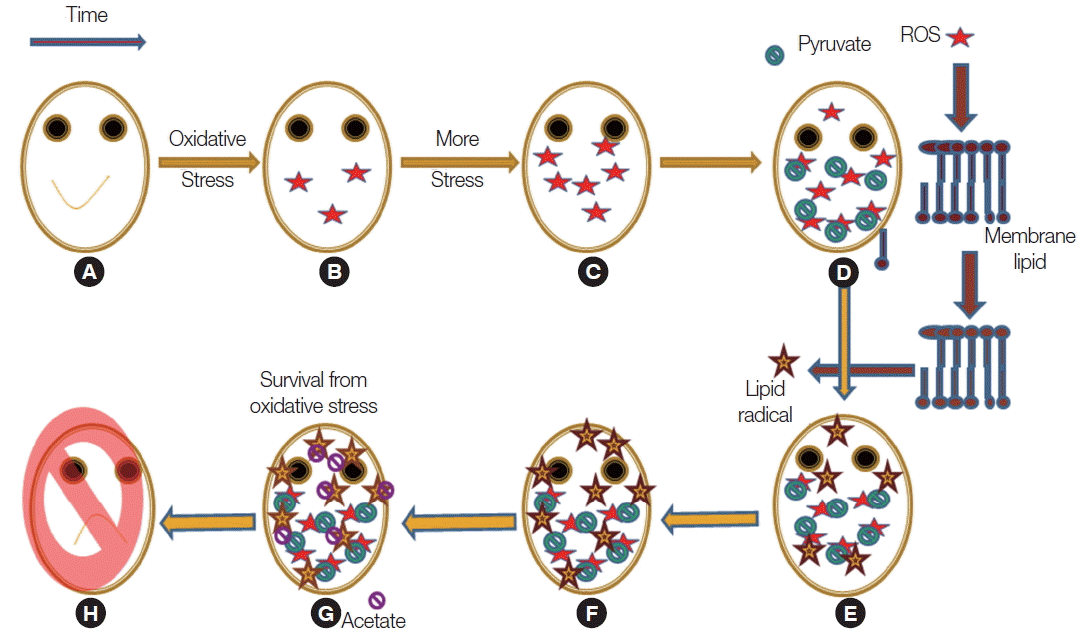
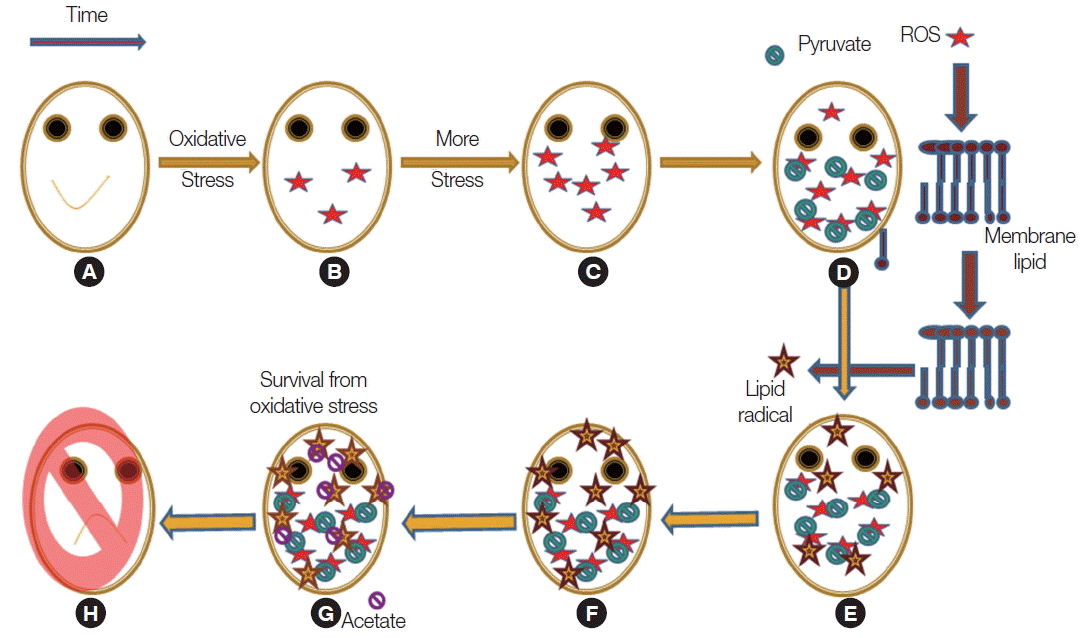
In trophozoites of
Giardia, hydroxyl and super oxide radicals are produced during cysteine-ascorbate deprivation (
Fig. 8). These radicals are spreads to the membrane and then initiate membrane lipid peroxidation. Thus, pyruvate, by eliminating hydroxyl and superoxide radical, prevented lipid peroxidation not lipid radicals. It was exciting to observe that acetate showed inhibition of lipid peroxidation, but the key mechanism still remains unknown.
In our present study, it was shown that intracellular pyruvate concentration was found to be increasing and it decreased after several hours. It can be explained by the inactivation of pyruvate: ferredoxin oxidoreductase, an enzyme which is sensitive to oxidative stress. However, it might also result from a metabolic need. Pyruvate concentration can also be increased by the regulation of pyruvate dikinase and malate dehydrogenase that Giardia can produce more pyruvate.
This study has revealed the dynamics of the transcriptional and metabolic regulatory networks during cysteine-ascorbate deprivation. Previously it was reported that transcriptional responses differ in
Giardia isolates of different genotypes and revealed higher basic transcript levels of antioxidant genes [
27].
However, the transcriptomic result shows that the oxidative stress management in
Giardia is not only regulated by some unconventional non-mitochondrial genes, but some metabolic genes play consequential role in the ROS regulation in cellular survival [
13,
28]. In the absence of any known machinery for oxidative stress management in
Giardia, this early divergent eukaryote combats the oxidative stress by modulating intracellular pyruvate concentration. In our previous study we have shown that the pyruvate protects
Giardia trophozoites from the exposure of hydrogen peroxide, a ROS generator used to mimic the gut environment in vitro [
29]. Here, we have tried to mimic the gut environment by generating oxidative stress by CAD medium. Although, transcriptomic data suggests that the stress survival in this case is mostly dependent on regulation of arginine deiminase as compared to malate dehydrogenase in case of H
2O
2. But, it is still observed that pyruvate remained the key metabolite and participated to reduce the oxidative stress by scavenging ROS in both the conditions.
This study has revealed the dynamics of the transcriptional and metabolic networks during cysteine-ascorbate deprivation. Previously it was reported that transcriptional responses differ in
Giardia isolates of different genotypes and revealed higher basic transcript levels of antioxidant genes [
27]. However, the transcriptomic results have shown that the oxidative regulation is not only controlled by some other genes, but also some metabolic genes have taken a significant role in ROS detoxification [
13,
28]. As
Giardia lacks conventional ROS detoxification enzyme cascades, oxidative stress management is controlled by modulating intracellular pyruvate concentration.
Notes
-
The authors have no financial or personal relationships with other people or organizations that could inappropriately influence or bias the content of this manuscript.
Supplementary materials
Supplementary Fig. 1.
H2DCFDA-loaded cells under confocal microscope after cysteine-ascorbate deprived (CAD) medium stress. Increases in fluorescence are representative of increase in the rate of oxidative species generated. Fluorescence was monitored from the suspension of live cells after the incubation under CAD medium in the absence and presence of pyruvate (2 mM) and acetate (2 mM). We used mannitol (2 mM) and ascorbate (2 mM) as a positive control and without metabolite as a negative control. Increases in fluorescence are representative of increase in the rate of oxidative species generated.
Supplementary Fig. 2.
Flow cytometric histograms of intracellular fluorescence intensity (488 nm excitation, 530 nm emission) in Giardia lamblia trophozoites under cysteine-ascorbate deprivation. Trophozoites were incubated at 35.5°C under CAD medium for 8 hr with increasing concentration of menadione in the absence and presence of pyruvate. Trophozoites were then washed and further incubated for 1 hr with or without pyruvate and replaced into the initial culture medium supplemented with the corresponding concentrations of pyruvate. Viability was estimated 24 hr later. The various cell populations reflect different treatments: (a) fluorescence arising from H2DCFDA-loaded without medium CAD treated cells, (b) H2DCFDA-loaded cells under CAD medium previously incubated with pyruvate (2 mM). (c) H2DCFDA-loaded cells under CAD medium previously incubated with pyruvate (0.5 mM), (d) H2DCFDA-loaded cells under CAD medium previously incubated with pyruvate (2 mM) in the presence of menadione (10 μM), (e) H2DCFDA-loaded cells under CAD medium previously incubated with pyruvate (2 mM) in the presence of menadione (30 μM), (f) H2DCFDA-loaded cells under CAD medium, (g) H2DCFDA-loaded cells under CAD medium previously incubated with menadione (10 μM), (h) H2DCFDA-loaded cells under CAD medium previously incubated with menadione (30 μM). All data were analyzed by WinMDI 2.9 software.
Supplementary Fig. 3.
Effect of pyruvate on DNA fragmentation. Electrophoretic analysis of DNA fragmentation on a 1.5% agarose for Giardia trophozoites treated with CAD medium for 8 hr in the absence and presence of pyruvate.
ACKNOWLEDGMENTS
This study was jointly supported by grants from the Okayama University Program of Founding Research Centre for Emerging and Re-emerging Infectious Disease, Ministry of Education, Culture, Sports, Science and Technology of Japan and National Institute of Infectious Diseases (NIID), Japan. The authors acknowledge Prof. Yushifomi Takeda and Prof. H. Wattanabe and Prof. Ichiro Kurane, NIID for their continuous constructive suggestions, support and critical review during this study. The authors also thank Indian Council of Medical Research, India for providing fellowship to DR during the study period and Mrs. Debarati Ganguly of Calcutta University for her help in proofreading this manuscript.
Fig. 1Pyruvate decreases the level of ROS in Giardia lamblia trophozoites under cysteine-ascorbate deprivation. Giardia trophozoites were incubated in cysteine-ascorbate deprived (CAD) medium and exposed to increasing doses of pyruvate (from 0–5 mM). Levels of ROS were estimated by spectrofluorometry using 2′, 7′-dichlorodihydrofluoresceine diacetate. Data are the mean±SEM of 3 independent experiments, each performed in triplicate. Pyr, Pyruvate; Man, Manitol. *P<0.05, compared with control; #P<0.001, compared with stressed sample.
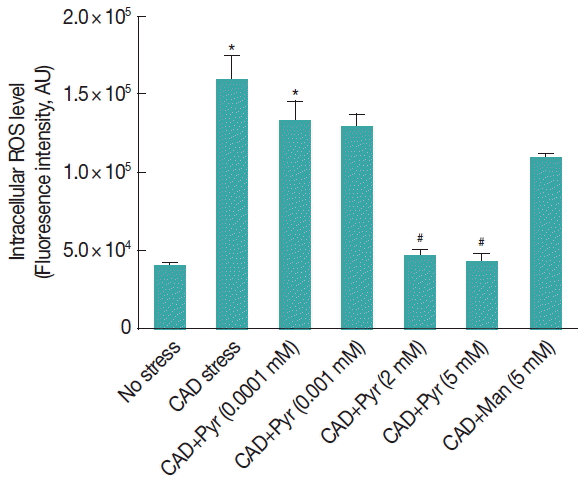
Fig. 2Pyruvate protects Giardia trophozoites from CAD medium induced cytoxicity. Trophozoites were incubated at 35.5°C upon CAD medium for 8 hr with increasing concentration of pyruvate. CAD medium-induced trophozoites were reseeded in fresh TYIS-33 medium and their viability was evaluated after 24 hr by using flowcytometry. Acetate (Ace) was shown not to decrease the rate of ROS generation in Giardia trophozoites. Results are expressed as the percentage of surviving trophozoites compared with control sample. Data are the mean±SEM of 3 independent experiments, each performed in triplicate. *P<0.05, compared with control; #P<0.001, compared with stressed sample.

Fig. 3ROS scavenging capacities and antioxidant properties of various α-ketoacids. Giardia trophozoites were preincubated for 30 min with a 2 mM concentration of each compound (Oxa=oxaloacetate, Mal=malate, Pyr=pyruvate, αKgl=α-ketoglutarate, Man=mannitol, Ace=acetate) and further incubated for 8 hr under CAD medium in their presence or absence. Viability was estimated 24 hr later. Data are the mean±SEM of 3 independent experiments, each performed in triplicate. *P<0.05, compared with control; #P<0.001, compared with stressed sample.
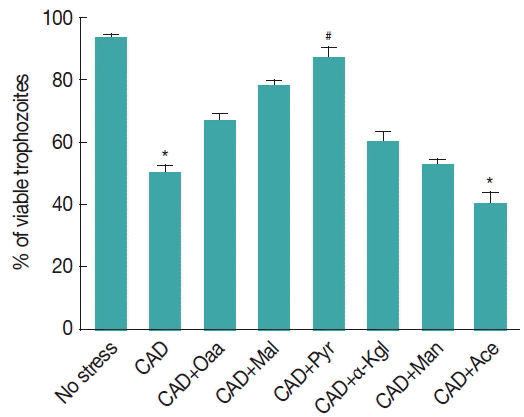
Fig. 4Effect of pyruvate on level of lipid peroxidation in cultured Giardia trophozoites upon CAD medium stress: MDA concentration in CAD medium stressed Giardia trophozoites after 8 hr incubation. Values are mean±SEM of 3 independent experiments, each performed in triplicate. *P<0.05, compared with control; #P<0.001, compared with stressed sample.
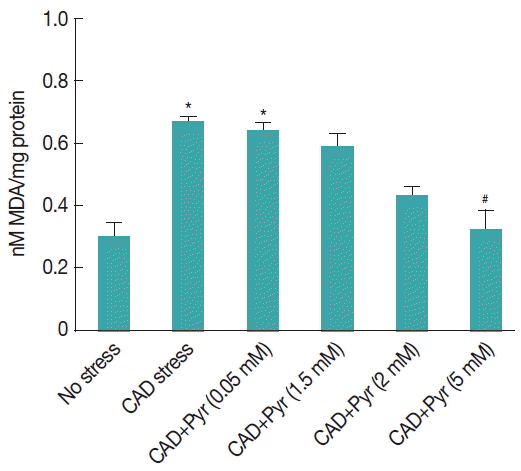
Fig. 5Intracellular pyruvate concentration in Giardia trophozoites during oxidative stress. Intracellular pyruvate content was measured in Giardia lamblia under CAD medium deprivation. The level of intracellular pyruvate was quantified every 2 hr interval. Values are means±SEM of 3 independent experiments, each performed in triplicate. *P<0.05, compared with control; ιP<0.001, compared with 4th hour stressed sample, τP<0.001, compared with 2nd hour stressed sample.
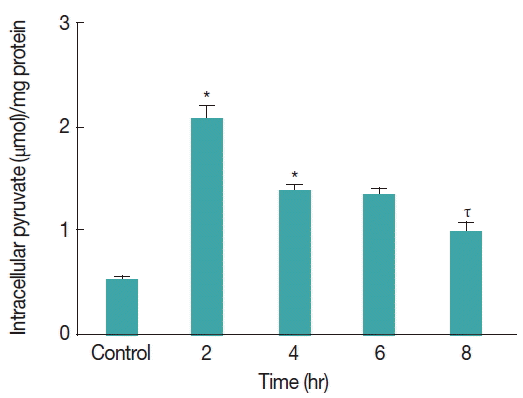
Fig. 6Quantification of viable cells. Counts of viable cells are given as percentages of the total number of cells. Trophozoites were incubated with CAD medium for 8 hr after that pyruvate was added and then further incubated for 30–120 min. Stress-induced trophozoites were reseeded in fresh TYI-S-33 medium supplemented with the corresponding concentration of pyruvate and their survival rate was evaluated after 24 hr by using flowcytometry. Results are expressed as the percentage of surviving trophozoites compared with control culture. Data are the mean±SEM of 3 independent experiments, each performed in triplicate. Pyr=pyruvate. *P<0.05, compared with control; #P<0.001, compared with stressed sample. Results are from 3 independent experiments, each performed thrice.
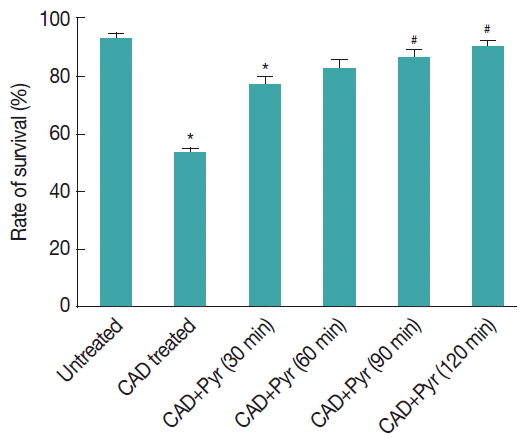
Fig. 7Effect of CAD medium on the expression of genes involved in pyruvate metabolism. Modulation of transcripts encoding enzymes involved in pyruvate metabolism. A gene expression (fold change) under CAD medium stress. Data are shown as fold change in relative expression compared with Actin on the basis of Comparative Ct (2−ΔΔCt) method. Values are shown as mean±SEM of 3 independent experiments, each performed in triplicate. (gene abbreviations used: metabolic genes: PFOR: pyruvate-ferredoxin oxidoreductade, MALDH: malate dehydrogenase, ARGD: arginine deiminase, PK: pyruvate kinase, PDK: pyruvate dikinase, ACS: acetyl coA synthase).
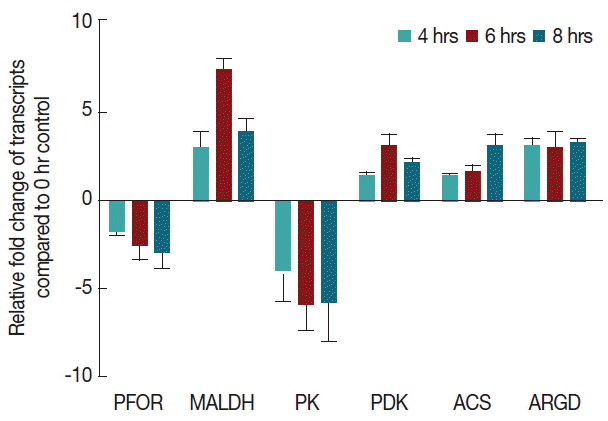
Fig. 8Schematic representation of the oxidative stress management in Giaradia. (A) Healthy cell. (B) Stressed cells accumulates intracellular ROS. (C) Concentration of intracellular ROS generation augmented. (D) Intracellular pyruvate concentration increases by the upregulation of malate dehydrogenase and pyruvate phosphate dikinase. (E) As ROS generation is a cyclic and also a spontaneous process it produces alcoxyl, peroxyl or lipid radicals by lipid peroxidation. (F) Lipid radicals are not scavenged by pyruvate so it produces more lipid radicals. (G) Intracellular acetate concentration is up-regulated as acetyl CoA synthase up-regulated. (H) On further stress, as pyruvate ferredoxin oxidoreductase is down-regulated so acetate production is inhibited and ultimate fate is death.
References
- 1. Adam RD. The biology of Giardia spp. Microbiol Rev 1991;55:706-732.
- 2. Adam RD. Biology of Giardia lamblia
. Clin Microbiol Rev 2001;14:447-475.
- 3. Lane S, Lloyd D. Current trends in research into the waterborne parasite Giardia
. Crit Rev Microbiol 2002;28:123-147.
- 4. Tak V, Mirdha BR, Yadav P, Vyas P, Makharia GK, Bhatnagar S. Molecular Characterization of Giardia intestinalis assemblages from human isolates at a tertiary care center of India. Ind J Med Micro 2014;32:19-25.
- 5. Farthing MJ. The molecular pathogenesis of giardiasis. J Pediatr Gastroenterol Nutr 1997;24:79-88.
- 6. Atkinson HJ. Respiration in Nematodes. In: Zuckerman BM, editor. Nematodes as Biological Models. 2:London, UK. Academic Press; 1980. p. 101-138.
- 7. Brown DM, Upcroft JA, Upcroft P. Free radical detoxification in Giardia duodenalis
. Mol Biochem Parasitol 1995;72:47-56.
- 8. Lujan HD, Byrd LG, Mowatt MR, Nash TE. Serum Cohn fraction IV-1 supports the growth of Giardia lamblia in vitro. Infect Immun 1994;62:4664-4666.
- 9. Gillin FD, Reiner DS, McCann PP. Inhibition of growth of Giardia lamblia by difluoromethylornithine, a specific inhibitor of polyamine biosynthesis. J Protozool 1984;31:161-613.
- 10. Brown DM, Upcroft JA, Upcroft P. Cysteine is the major low-molecular weight thiol in Giardia duodenalis
. Mol Biochem Parasitol 1993;61:155-158.
- 11. Tekwani BL, Mehlotra RK. Molecular basis of defence against oxidative stress in Entamoeba histolytica and Giardia lamblia
. Microbes Infect 1999;1:385-394.
- 12. Ghosh E, Ghosh A, Ghosh AN, Nozaki T, Ganguly S. Oxidative stress-induced cell cycle blockage and a protease-independent programmed cell death in microaerophilic Giardia lamblia
. Drug Des Devel Ther 2009;3:103-110.
- 13. Raj D, Ghosh E, Mukherjee AK, Nozaki T, Ganguly S. Differential gene expression in Giardia lamblia under oxidative stress: significance in eukaryotic evolution. Gene 2014;535:131-139.
- 14. Paget TA, Kelly ML, Jarrol EL, Lindmark DG, Lloyd D. The effects of oxygen on fermentation in Giardia lamblia
. Mol Biochem Parasitol 1993;57:65-72.
- 15. Mead JF. Free Radical Mechanisms of Lipid Damage and Consequences for Cellular Membranes. In: Pryor WA, editor. Free Radicals in Biology. New York, USA. Academic Press; 1976. p. 51-68.
- 16. Schuessel K, Frey C, Jourdan C, Keil U, Weber CC, Müller-Spahn F, Müller WE, Eckert A. Aging sensitizes toward ROS formation and lipid peroxidation in PSIM146L transgenic mice. Free Radic Biol Med 2006;40:850-862.
- 17. Bar-Or D, Rael LT, Lau EP, Rao NK, Thomas GW, Winkler JV, Yukl RL, Kingston RG, Curtis CG. An analog of the human albumin N-terminus (Asp-AlaHis-Lys) prevents formation of copper-induced reactive oxygen species. Biochem Biophys Res Commun 2001;284:856-862.
- 18. Evans PJ, Gallesi D, Mathieu C, Hernandez MJ, Felipe M, Halliwell B, Puppo A. Oxidative stress occurs during soybean nodule senescence. Planta 1999;208:73-79.
- 19. Rael LT, Thomas GW, Craun ML, Curtis CG, Bar-Or R, Bar-Or D. Lipid peroxidation and the thiobarbituric acid assay: standardization of the assay when using saturated and unsaturated fatty acids. J Biochem Mol Biol 2004;37:749-752.
- 20. Nandi N, Sen A, Banerjee R, Kumar S, Kumar V, Ghosh AN, Das P. Hydrogen peroxide induces apoptosis-like death in Entamoeba histolytica trophozoites. Microbiology 2010;156:1926-1941.
- 21. Andrae U, Singh J, Ziegler-Skylakakis K. Pyruvate and related alpha-ketoacids protect mammalian cells in culture against hydrogen peroxide-induced cytotoxicity. Toxicol Lett 1985;28:93-98.
- 22. Crestanello JA, Kamelgard J, Whitman GJ. The cumulative nature of pyruvate’s dual mechanism for myocardial protection. J Surg Res 1995;59:198-204.
- 23. Holleman AF. Notice sur l’action de l’eau oxygénée sur les acides αcétoniques et sur les dicétones 1.2. Recl Trav Chim Pays Bas 1904;23:169-171. (in French).
- 24. Biagini GA, Park JH, Lloyd D, Edwards MR. The antioxidant potential of pyruvate in the amitochondriate diplomonads Giardia intestinalis and Hexamita inflata
. Microbiology 2001;147:3359-3365.
- 25. Herbener GH. A morphometric study of age-dependent changes in mitochondrial population of mouse liver and heart. J Gerontol 1976;31:8-12.
- 26. Vorbeck ML, Martin AP, Park JK, Townsend JF. Aging related decrease in hepatic cytochrome oxidase of the Fischer 344 rat. Arch Biochem Biophys 1982;214:67-79.
- 27. Ma’ayeh SY, Knörr L, Svärd SG. Transcriptional profiling of Giardia intestinalis in response to oxidative stress. Int J Parasitol 2015;45:925-938.
- 28. Ganguly S, Raj D. Giardia and Giardiasis: an Overview of Recent Developments. The Handbook of Microbial Bioresources. In: Gupta VK, Sharma GD, Tuohy MG, Gaur R, editors. The Handbook of Microbial Bioresources. London, UK. CABI Wallingford; 2016. p. 211-227.
- 29. Raj D, Saini P, Nozaki T, Ganguly S. Involvement of pyruvate on oxidative stress management in the microaerophilic protozoan parasite Giardia lamblia
. Int J Adv Res 2015;4:1148-1166.