Abstract
To evaluate whether iron concentration in TYM medium influence on hydrogenosomal enzyme gene expression and hydrogenosomal membrane potential of Trichomonas vaginalis, trophozoites were cultivated in iron-depleted, normal and iron-supplemented TYM media. The mRNA of hydrogenosomal enzymes, such as pyruvate ferredoxin oxidoreductase (PFOR), hydrogenase, ferredoxin and malic enzyme, was increased with iron concentrations in T. vaginalis culture media, measured by RT-PCR. Hydrogenosomal membrane potentials measured with DiOC6 also showed similar tendency, e.g. T. vaginalis cultivated in iron-depleted and iron-supplemented media for 3 days showed a significantly reduced and enhanced hydrogenosomal membrane potential compared with that of normal TYM media, respectively. Therefore, it is suggested that iron may regulate hydrogenosomal activity through hydrogenosomal enzyme expression and hydrogenosomal membrane potential.
-
Key words: Trichomonas vaginalis, hydrogenosomal enzyme, gene expression>, hydrogenosomal membrane potential, iron, DiOC6, PFOR, hydrogenase, malic enzyme, ferredoxin
Trichomonas vaginalis is a common parasite found in females, worldwide (
Beaver et al., 1984). Recent data on the prevalence of trichomoniasis in Korea showed that 10.4% (17/163) of females who visited Hanyang University Kuri Hospital in Kyonggi-do and 7.6% (478/6,262) who visited Yonsei University Wonju Hospital in Kangwon-do, were found to carry
T. vaginalis as diagnosed by PCR or a wet mount (
Choi et al., 1996;
Ryu et al., 1999).
Hydrogenosomes, an unusual organelle found in the amitochondriate eukaryote,
T. vaginalis, play an essential role in the drug susceptibility of
T. vaginalis (
Quon et al., 1992). Trichomoniasis is treated with metronidazole, derivatives of 5-nitroimidazole which frequently used to treat infections caused by a number of anaerobic pathogens. Metronidazole is administered in an inactive form and is reduced to its cytotoxic form within the hydrogenosomes. Hydrogenosomal proteins such as pyruvate ferredoxin oxidoreductase (PFOR), hydrogenase, ferredoxin and malic enzyme, play a critical role in the reductive activation of metronidazole (
Müller, 1993).
Iron is an essential nutrient for the growth of many pathogens, including
T. vaginalis (
Griffith, 1985;
Gorrell, 1985). Several reports have recently shown that iron can regulate a variety of trichomonal properties such as overall adhesin synthesis, cytoadherence level, gene expression of surface immunogens and resistancy to complement lysis provided by immune evasion strategy of the
T. vaginalis (
Lehker et al., 1991;
Lehker and Alderete, 1992;
Alderete et al., 1995a;
Alderete et al., 1995b). Our previous data have shown that the virulence of
T. vaginalis is reduced under iron-depleted conditions (
Ryu et al., 2001). However, it is not known whether iron also affects hydrogenosomal enzyme gene expression. In this study, we examined whether iron concentration in a complex Trypticase-yeast extract-maltose (TYM) medium influence on hydrogenosomal enzyme gene expression and hydrogenosomal membrane potential of
T. vaginalis.
Trichomonads were grown in TYM medium, supplemented with 10% heat-inactivated horse serum (
Diamond, 1957). The iron-deficient medium was prepared by adding 2,2'-dipyridyl (Sigma, St. Louis, Missouri, USA) at 100 µM final concentration, whereas the iron-supplemented medium was prepared with the addition of ferrous sulfate at a final concentration of 360 µM (
Ryu et al., 2001). Iron concentrations of iron-deficient, normal, and iron-supplemented media were 1.1 µM, 34.8 µM, and 393 µM, respectively, as determined by an automatic chemical analyzer (Hitachi Inc, Tokyo, Japan). The 2,2'-dipyridyl, used for iron-deficient conditions, is chemically recognized as a specific Fe
++ chelator, with very little reactivity with Fe
+++. Because of its small size, hydrophobic nature, and its rapid permeation into cells, dipyridyl acts intracelluarly (
Horky et al., 1998).
Early log-phase trophozoites (2 × 10
5) were inoculated into 5 ml of normal TYM, iron-deficient TYM, and iron-supplemented TYM media. The next day, the same number of trichomonads were transferred into fresh media and grown for another 24 hr. Subculture of trophozoites was done in same manner for 4 days (
Ryu et al., 2001).
For RT-PCR, total RNA was isolated using the Trizol reagent (Invitrogen, Carlsbad, California, USA). Total RNA from
T. vaginalis grown in iron depleted, normal and iron enriched media for 4 days were reverse transcribed with oligo (dT) 15 primer using MMLV reverse transcriptase (Invitrogen). PCR amplification of cDNA was carried out using gene-specific primers (
Table 1). The PCR reaction consisted of 1 unit of Taq polymerase (Takara, Otsu, Japan), 2 µl of 10 mM dNTP mix, 2 µl of 10× PCR buffer, 5.2 µl of 5 M betaine, 0.5 µl of DMSO, 20 pM of forward and reverse primers, and 2.0 µl of template cDNA in a total volume of 20 µl. Cycling was performed using the conditions of initial DNA denaturation at 94℃ for 5 min and 25 rounds of denaturation (94℃ for 1 min), annealing (65℃ for 1 min), and extension (72℃ for 2 min). β-tubulin was amplified as an internal control. After 25 cycles of amplification by PCR, 10 µl of the PCR products were electrophoresed in 1% agarose gel containing 0.5 µg/ml ethidium bromide, and photographed under ultraviolet light. The band intensity was quantified using the Quantity One program (Bio-Rad, Hercules, California, USA).
Next, we measured hydrogenosomal membrane potential of
T. vaginalis cultivated in different iron concentration of TYM media by FACS analysis after DiOC
6 staining.
T. vaginalis cultured in iron-deficient, normal, and iron-supplemented media were washed with cold PBS, after which cells (5 × 10
5/ml) were incubated with DiOC
6 (40 nM in PBS) for 15 min at 37℃ (
Chose et al., 2002). Flow cytometric analysis for fluorescent intensity was performed on at least 5,000 cells from each sample with a FACSCalibur with CellQuest pro software (BD Bioscience, Germany). The hydrogenosomal enzyme expressions and hydrogenosomal membrane potentials of
T. vaginalis cultivated in iron supplemented and iron-depleted media were compared with those of normal TYM media.
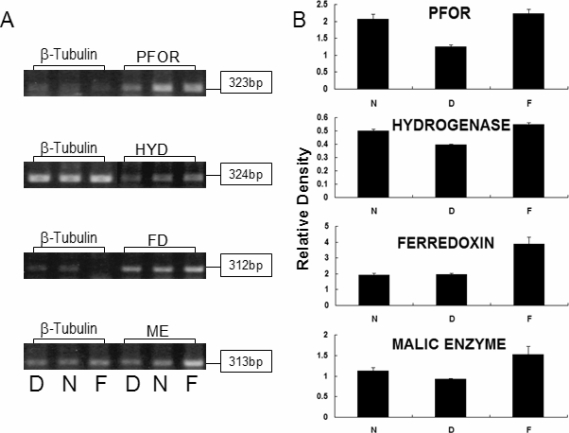
As a result of RT-PCR, mRNA levels of PFOR, hydrogenase and malic enzyme were reduced 61-83% in iron-depleted media although ferredoxin mRNA showed same level as trophozoites in normal media (
Fig. 1). Moreover, trophozoites grown in iron-supplemented TYM media had higher gene expression of 110-120%. Therefore, the gene expressions of hydrogenosomal enzyme such as PFOR, hydrogenase, ferredoxin and malic enzyme were thought to be increase with iron concentration in
T. vaginalis culture media.
We identified that trophozoites in iron-rich media showed increased band intensity (128 kDa) with anti-PFOR antibody compared with that of normal media by Western blot (data not shown). Therefore, it is assumed that increased PFOR mRNA expression of trophozoites in iron-supplemented media may be correlated with increased PFOR protein. The present study supports the results of Gorrell (1985), which showed that T. vaginalis grown in iron-rich medium increased concentrations of iron-sulfur proteins, including ferredoxin and PFOR.
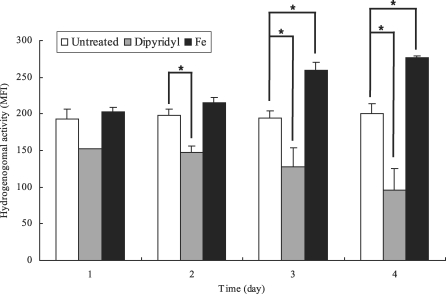
Next, hydrogenosomal membrane potential of
T. vaginalis in iron-depleted media was decreased with cultivation time. In contrast, trophozoites grown in iron-supplemented media showed increased hydrogenosomal potentials in time dependent manner. After 3 days of incubation,
T. vaginalis in iron-depleted and iron-supplemented media exhibited significant reduced and enhanced hydrogenosomal membrane potential compared with that of normal TYM media, respectively. Hydrogenosomal membrane potentials of trophozoites grown in each media for 4 days showed 50% lower level in iron-deficient media and 143% higher fluorescent intensity in iron-rich media (
Fig. 2).
T. vaginalis lacks morphologically recognizable mitochondria but contains hydrogenosomes which is homologous to mitochondria of eukaryotes (
Müller, 1993). These characteristic organelles are predominantly spherical or somewhat elongated structures, about 0.5 µm in diameter. Hydrogenosome had been recognized by light microscopists for a long time, as paraxostylar and paracostal granules. They were named hydrogenosomes because they produce molecular hydrogen as a metabolic end product. The hydrogenosomes represent an integral part of the energy metabolism of
T. vaginalis because certain characteristic steps of carbohydrate metabolism linked to substrate level phosphorylation occur in this organelle.
T. vaginalis have been developed resistance to metronidazole in a multistep process based on gradual decrease and eventual loss of activities of the hydrogenosomal protein such as PFOR, hydrogenase, ferredoxin and malic enzyme involved in drug-activating pathway, by prolonged exposure to increasing concentrations of metronidazole (
Kulda et al., 1993;
Rasoloson et al., 2002). Quon et al. (
1992) confirmed that metronidazole-resistant
T. vaginalis strains isolated from trichomoniasis patients showed reduced transcriptions of the ferredoxin gene. The decreased hydrogenosomal enzyme expression of
T. vaginalis cultivated in iron-depleted media in the present study seems like to that of metronidazole-resistant strain artificially developed with increasing concentration of metronidazole.
DiOC
6 has been generally used for detection of apoptotic cells by measurement of decreased mitochondrial membrane potentials (
Kim et al., 2000;
Kang et al., 2006) although this drug was also used for observation of mitochondria, endoplasmic reticulum and sarcoplasmic reticulum in eukaryotes (
Habicht and Brune, 1980;
Johnson et al., 1981;
Terasaki et al., 1986). Chose et al. (
2002) applied DiOC
6 marker for measuring dissipation of hydrogenosomal membrane potential of amitochondriate,
T. vaginalis, pretreated with toxic agents in order to determine apoptosis of trophozoites, and observed a weak but significant membrane potential dissipation in pro-apoptotic drugs-treated trichomonads.
In contrast, in this study, we tried to use DiOC6 for measuring of hydrogenosomal membrane potentials of live trichomonads (showing 100% viability in trypan blue exclusion test) cultivated in different iron condition. The hydrogenosomal membrane potentials of trichomonads increased with increment of iron contents in TYM media, and this result had similar tendency with results of the hydrogenosomal enzyme mRNA analysis.
Until now, reports on relation between hydrogenosomal membrane potentials and hydrogenosomal enzyme expression could not be found. Viable trichomonads (from normal and iron-rich media) pretreated with anti-PFOR antibody showed decreased hydrogenosomal membrane potentials with DiOC6 staining (data not shown).
In our previous report, the virulence of
T. vaginalis in iron depleted condition is described to be reduced through decrease in subcutaneous abscess formation, cytoadherence, cytotoxicity, and proteinase activity (
Ryu et al., 2001). Hydrogenosomal enzyme expression and hydrogenosomal membrane potential were also decreased under the same iron-depleted media described by Ryu et al. (
2001). Taking this altogether, it is expected that iron may regulate the virulence of
T. vaginalis via hydrogenosomal activity. In further study, it should be evaluated whether hydrogenosomal activity under different iron condition may influence on virulence of
T. vaginalis.
References
Fig. 1RT-PCR of hydrogenosomal enzymes of T. vaginalis cultivated in iron-depleted, normal and iron-supplemented TYM media. A. PCR results of cDNA from T. vaginalis cultivated from iron-depleted TYM containing 100 µM 2,2'-dipyridyl (D), from normal TYM (N), and from iron-supplemented TYM media containing 360 µM ferrous sulfate (F). B. Density of PCR band from amplified hydrogenosomal enzyme of T. vaginalis was measured by Quantity One (version 4.6.2, BioRad, USA) and compensated by the density of PCR band from amplified β-tubulin of the same T. vaginalis. The results were marked as relative density. PFOR = pyruvate ferredoxin oxidoreductase, HYD = hydrogenase, FD = ferredoxin, ME = malic enzyme.
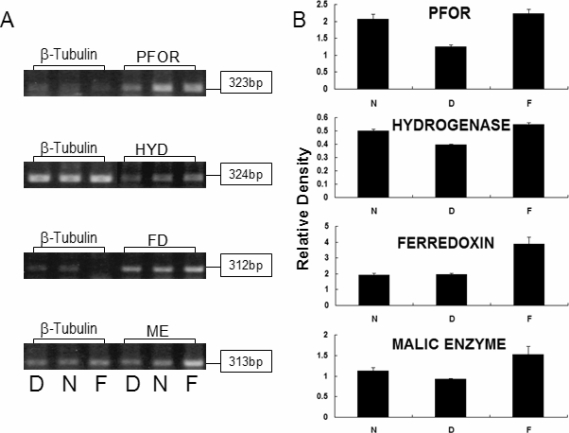
Fig. 2Mean fluorescent intensities (MFI) of hydrogenosomal membrane potentials of Trichomonas vaginalis cultivated in normal (untreated, □), iron-depleted (Dipyridyl, ▒) and iron-supplmented (Fe, ▪) TYM media from one to four day of infection, measured with flow cytometry after DiOC6 staining. The data are expressed as the means ± SE of 3 separate experiments. *P value < 0.05.
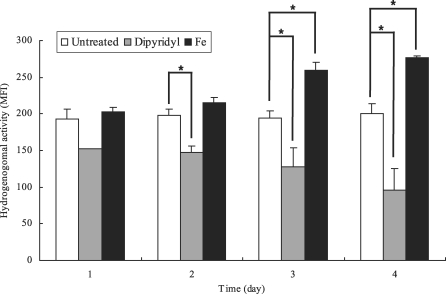
Table 1.Primers for RT-PCR of hydrogenosomal enzymes from Trichomonas vaginalis
Table 1.
|
Hydrogenosomal enzyme |
Primer sequence |
|
Ferredoxin (genebank ID= 39939566) CDS-nta) 1 |
5´ ATGCTCTCTCAGTGCTCTCCTC 3´ (forward) |
|
Ferredoxin (genebank ID= 39939566) CDS-nta) 312 |
5´ TTAGACCTCGAATGTAGCACCG 3´ (reverse) |
|
Hydrogenase (genebank ID= 1171116) CDS-nta) 778 |
5´ GGAAAGCAAGAGACAGGTGC 3´ (forward) |
|
Hydrogenase (genebank ID= 1171116) CDS-nta) 1101 |
5´ TGCATTCTTTATGCCGTGAG 3´ (reverse) |
|
Malic enzyme (genebank ID= 33243007) CDS-nta) 144 |
5´ CCTCGAGATCCAGAAGAACG 3´ (forward) |
|
Malic enzyme (genebank ID= 33243007) CDS-nta) 456 |
5´ TTGGAGACGCTTCTCGATTT 3´ (reverse) |
|
Pyruvate:ferredoxin oxidoreductase (genebank ID= 622957) CDS-nta) 3015 |
5´ TGCTGCTGGTTACACAAAGG 3´ (forward) |
|
Pyruvate:ferredoxin oxidoreductase (genebank ID= 622957) CDS-nta) 3337 |
5´ GGTCGAGGTTGTAGTCTGGC 3´ (reverse) |
|
Beta-tubulin (genebank ID= 797282) CDS-nta) 480 |
5´ CCCAGATCGTATCCTCTCCA 3´ (forward) |
|
Beta-tubulin (genebank ID= 797282) CDS-nta) 791 |
5´ AGACGTGGGAATGGAACAAG 3´ (reverse) |