Abstract
Parvatrema chaii n. sp. (Digenea: Gymnophallidae) is described using the worms recovered from experimentally infected mice in Korea. The metacercariae were collected from surf-clams, Mactra veneriformis, from a tidal flat in Sochon-gun, Chungchongnam-do. The metacercariae were elliptical (0.262 × 0.132 mm), and the genital pore had an anterior arch of 16-17 sensory papillae in scanning electron microscopic view. Adult worms were ovoid to foliate (0.275-0.303 by 0.140-0.150 mm), and their characteristic features included the presence of lateral lips, short esophagus, genital pore located some distance anterior to the ventral sucker, club-shaped seminal vesicle, a compact to slightly lobed vitellarium, elliptical eggs (0.018-0.020 by 0.010-0.013 mm), and absence of the ventral pit. This gymnophallid is classified as a member of the genus Parvatrema because of the location of the wide genital pore some distance from the ventral sucker, and the absence of the ventral pit. It differs from previously reported Parvatrema species, including the type species, P. borinquenae. In particular, the morphologies of the vitellarium and the genital pore with an anterior arch of 16-17 sensory papillae are unique features. Therefore, we propose it as a new species, Parvatrema chaii n. sp. (Digenea: Gymnophallidae).
-
Key words: Parvatrema chaii n. sp., gymnophallid, mouse, surf-clam, Mactra veneriformis
INTRODUCTION
Gymnophallid flukes are exclusively marine parasites which commonly utilize bivalve mollusks and rarely utilize gastropods and polychaetes as intermediate hosts. Shore birds play the role of the definitive host with the exception of
Gymnophalloides seoi (
Lee et al., 1993;
Ching, 1995). The metacercariae of
Parvatrema timondavidi were found in
Tapes philippinarum, a marine clam species (
Yu et al., 1993), and those of
Gymnophalloides seoi, the only human-infecting species so far known, were detected in oysters,
Crassostrea gigas, in Korea (
Lee et al., 1995). In the present study, we detected gymnophallid metacercariae in surf-clams,
Mactra veneriformis, and recovered adult worms from experimental mice that had been infected with these metacercariae. On the basis of the unique morphological characteristics of both stages, as observed via light microscopy and scanning electron microscopy (SEM), we intend to propose it as a new species,
Parvatrema chaii n. sp. (Digenea: Gymnophallidae).
MATERIALS AND METHODS
A total of 95 surf-clams,
M. veneriformis (
Fig. 1), were collected from a tidal flat located in Sochon-gun, Chungchongnam-do, Republic of Korea. The clams were individually examined via pepsin-HCl digestion in order to determine the infection status of the gymnophallid metacercariae. In order to acquire adult worms, 4 mice (ICR strain) were orally infected with 300 metacercariae each, via a gavage needle. The mice were sacrificed by cervical dislocation 1 week after infection. Their small intestines were resected and divided into 3 equal pieces, then opened longitudinally. The adult worms were then harvested from the intestinal contents of each of the intestinal pieces under a stereomicroscope.
The metacercariae and adult worms were fixed using 10% neutral buffered formalin under a coverslip pressure, stained with Semichon's acetocarmine, and observed under a light microscope. Some of the metacercariae were prepared for SEM, and were observed with a SEM (ISI-Korea DS-130C) at an acceleration voltage of 10 kV.
RESULTS
Infection status of the clam with the metacercariae
Out of 95 clams, 94 (98.9%) were infected with gymnophallid metacercariae (av. 217; 6-1,737 in range). The metacercarial infection status is shown in
Table 1.
A total of 302 adult flukes (25.2%) were recovered from 4 mice infected with 300 metacercariae each 1 week prior to the worm recovery. The recovery rate of worms from portions of the small intestine is depicted in
Table 2.
DESCRIPTION
Parvatrema chaii n. sp. (Figs. 2-8)
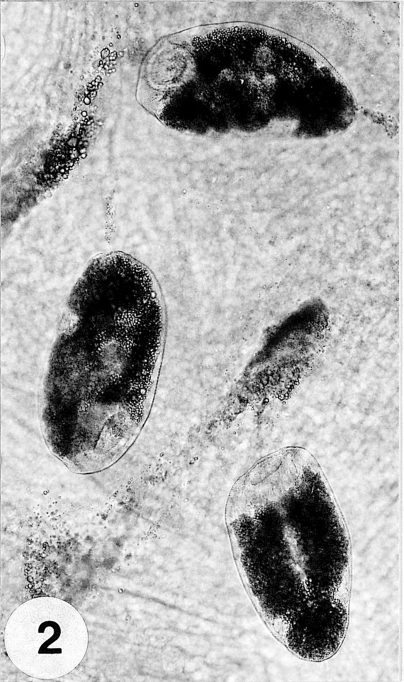
Metacercariae: Metacercariae were collected from the tissue between the mantle and shell (
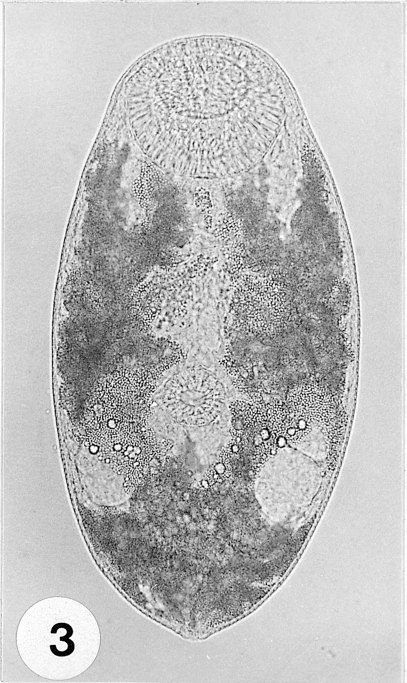
Fig. 2), and were elliptical, rounded anteriorly, slightly pointed posteriorly, 233-300 (262 ± 23.0) µm long, 125-140 (132 ± 5.0) µm wide at midbody. Oral sucker subterminal, large and muscular, 58-68 (63 ± 3.0) × 60-73 (69 ± 3.0) µm in size, with prominent lateral projections on each side, ratio to body length 1: 3.97. Prepharynx not seen. Pharynx well developed, muscular, 15-25 (20 ± 3.0) × 23-25 (24 ± 1.0) µm in size. Esophagus short. Ceca short, saccate, usually ending before mid-body. Ventral sucker round, 23-25 (25 ± 1.0) × 25-28 (26 ± 1.0) µm in size, located 2/5 of body length from the posterior end, sucker width ratio 1: 0.38 on average. Testes round to elliptical, 30-43 (34 ± 4.0) × 25-30 (27 ± 2.0) µm and 25-38 (32 ± 3.0) × 23-28 (25 ± 1.0) µm in size, located at both lateral sides, 1/4 body length from the posterior end. Ovary oval, 25-33 (29 ± 2.0) × 20-25 (23 ± 2.0) µm in size, located just anterior to the right testis. Vitellarium oblong, 18-28 (23 ± 3.0) × 10-18 (14 ± 2.0) µm in size. Excretory vesicle V-shaped, with anterior arms at the level of the pharynx (
Figs. 3,
4).
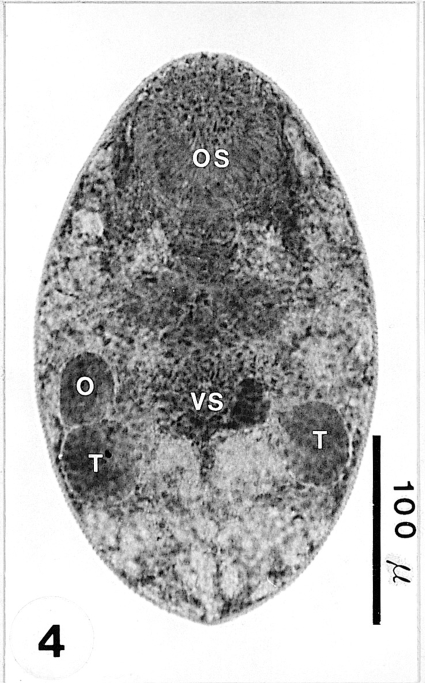
Adults: Adult flukes ovoid to foliate with anterior end more rounded than the posterior end, 275-303 (288 ± 10.0) µm long, 140-150 (147 ± 4.0) µm wide at the midbody. Oral sucker subterminal, large and muscular, 65-73 (69 ± 2.0) × 73-75 (75 ± 1.0) µm in size, with prominent lateral projections on each side, ratio to body length 1: 4.0. Prepharynx not seen. Pharynx rounded and muscular, 20-25 (22 ± 2.0) × 23-25 (23 ± 1.0) µm in size, leading to short esophagus. Ceca short and saccate. Ventral pit absent. Genital pore located some distance anterior to the ventral sucker. Ventral sucker round, 25-28 (26 ± 1.0) × 25-26 (26 ± 1.0) µm in size, located in posterior 2/5 of body length from the posterior end, sucker width ratio 1: 0.35. Testes ovoid, 43-53 (48 ± 4.0) × 28-33 (30 ± 1.0) µm and 40-50 (47 ± 4.0) × 23-33 (29 ± 4.0) µm in size, located somewhat diagonally to each other at the level of the mid hindbody. Seminal vesicle club-shaped, 30-40 (35 ± 3.0) × 25-38 (30 ± 4.0) µm in size. Ovary ovoid, 38-58 (50 ± 6.0) × 25-40 (32 ± 4.0) µm in size, located far anteriorly at the level of ceca. Vitellarium oblong, compact to slightly lobed, 30-50 (42 ± 5.0) × 23-28 (25 ± 1.0) µm in size. Uterus extending into forebody at the level of the pharynx. Eggs numerous, 18-20 (19.5) × 10-13 (11) µm in size. Excretory vesicle V-shaped, with anterior arms at the level of the pharynx (
Figs. 5,
6).
Taxonomic summary
Type host: Mactra veneriformis (intermediate) & ICR mouse (definitive).
Location: Small intestine.
Locality: Sochon-gun, Chungchongnam-do, Republic of Korea.
Specimens deposited: Holotype, GNU (Gyeongsang National University) Helm. Coll. no. 9701; paratypes, USNM Helm Coll. (Beltsville) no. 097225 & 097226.
Etymology: The specific name is in honor of Professor Jong-Yil Chai, Department of Parasitology and Tropical Medicine, Seoul National University College of Medicine, who has dedicated all his passion to the study of parasitology in Korea.
SEM findings of P. chaii metacercariae
Metacercariae were ovoid and slightly curved ventrally. The oral sucker was approximately 2.5 times larger than the ventral sucker (
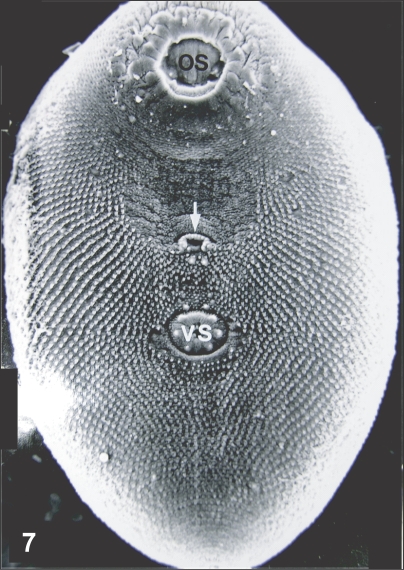
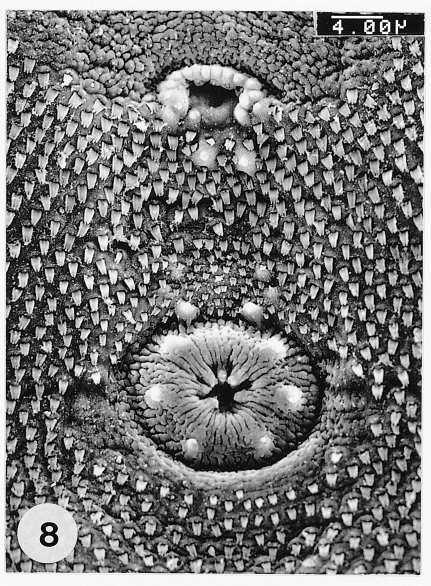
Fig. 7). The genital pore was located some distance anterior to the ventral sucker and had an anterior arch of 16-17 sensory papillae. The ventral sucker was elliptical, had a radially wrinkled tegument and 6 type I papillae symmetrically distributed on its rim (
Fig. 8).
DISCUSSION
The new trematode has been assigned as a member of the genus
Parvatrema in consideration of the location of the wide genital pore some distance from the ventral sucker, as well as the absence of a ventral pit. It appears to have a unique arrangement of sensory papillae surrounding the genital pore. This characteristic is difficult to see via light microscopy, but can be distinctively seen via SEM (
Pekkarinen and Ching, 1994;
Yu et al., 1994).
Three species previously described to have a single vitellarium, namely
P. timondavidi,
P. borinquenae, and
P. bushi, are comparable with the new species. However,
P. timondavidii has an ovoid body, an oral sucker only twice as large as the ventral sucker, and a highly lobed-vitellarium. Their egg sizes are larger than those of
P. chaii (
Yu et al., 1993). The type species,
P. borinquenae, has been reported as a metacercaria in the style sac of a gastropod mollusk, and as an adult in charadriiform birds (
Cable, 1953). The egg sizes and pharyngeal length are larger than those observed in
P. chaii (
Cable, 1953). The new species also differs from
P. bushi in its body shape, the possession of a wide genital pore with no sensory papillae, and the shape of the vitellarium, which is compact and rounded (
Ching, 1995).
The location of the ovary in the far right of the forebody in the new species is similar to that of
Meiogymnophallus macrostoma, which was reported in shore birds in Korea. However,
M. macrostoma differs from the new species with regard to the presence of very large paired vitellaria, and the location of the genital pore almost on the ventral sucker (
Yamaguti, 1939). The new species is also different from all other previously reported
Parvatrema species. Therefore, we propose it as a new species,
Parvatrema chaii n. sp. (Digenea: Gymnophallidae).
References
Fig. 1
Mactra veneriformis, the second intermediate host of Parvatrema chaii, collected from a tidal flat in Sochon-gun, Chungchongnam-do, Korea.

Fig. 2Three metacercariae parasitized on the mantle of the surf-clam, Mactra veneriformis.
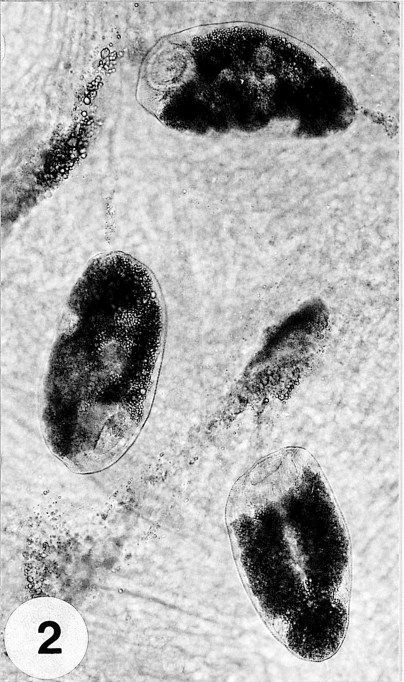
Fig. 3A fresh metacercaria isolated from a surf-clam, not encysted and having a Y-shaped excretory bladder.
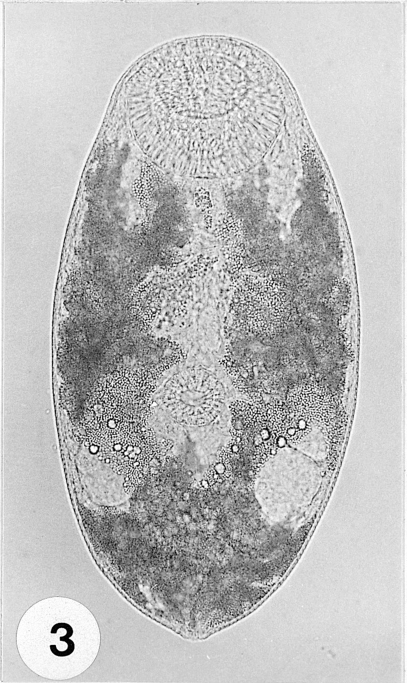
Fig. 4A metacercaria stained with Semichon's acetocarmine. Large oral sucker (OS), smaller ventral sucker (VS), elliptical ovary (O) and 2 round testes (T) can be distinctly observed.
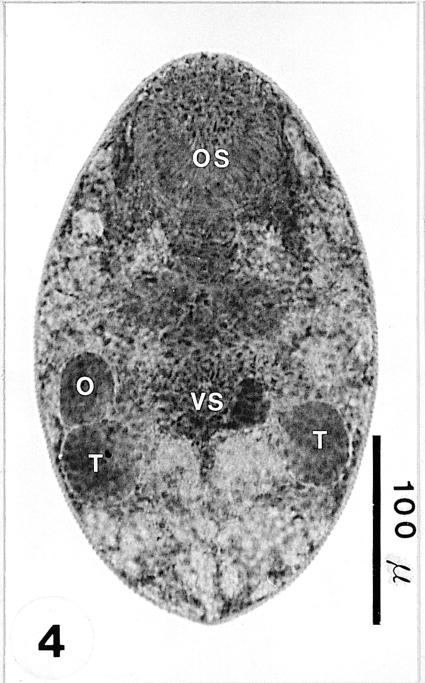
Fig. 5An adult recovered from an experimental mouse 1 week post-infection (PI), and stained with Semichon's acetocarmine. Genital organs, i.e ovary (O), genital pore (GP), seminal vesicle (SV), vitellarium (V), and 2 testes (T) can be clearly observed.

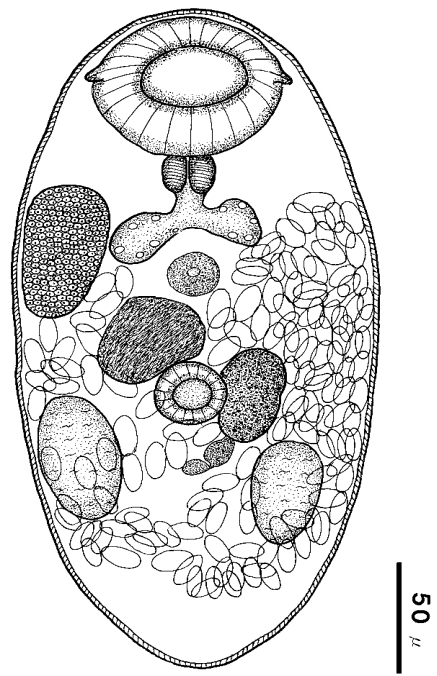
Fig. 6Drawing of Parvatrema chaii n. sp. (holotype), ventral view, recovered from an experimental mouse.

Fig. 7Scanning electron microscopic (SEM) view of P. chaii metacercariae. Large oral sucker (OS), genital pore (arrow) and small ventral sucker (VS) are characteristic.

Fig. 8SEM view of the genital pore and ventral sucker in the ventro-median portion of P. chaii metacercariae. The genital pore uniquely has an anterior arch of 16-17 sensory papillae.
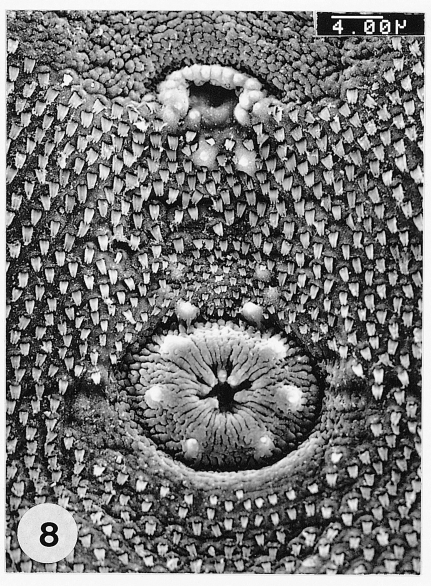
Table 1.Infection status of Parvatrema chaii metacercariae in Mactra veneriformis
Table 1.
|
Weight of clam (g) |
No. of clams exam. |
No. (%) clams posit. |
No. of metacercariae detected
|
|
Total |
Range |
Average ± SD |
|
Below 10 |
10 |
10 (100.0) |
513 |
6 - 182 |
51 ± 60 |
|
10.1 ~ 15.0 |
22 |
21 (95.5) |
2,546 |
11 - 404 |
121 ± 114 |
|
15.1 ~ 20.0 |
38 |
38 (100.0) |
8,630 |
6 -1,737 |
227 ± 313 |
|
Over 20.1 |
25 |
25 (100.0) |
8,743 |
13 -1,203 |
350 ± 315 |
|
Total |
95 |
94 (98.9) |
20,432 |
6 -1,737 |
217 ± 284 |
Table 2.Recovery of Parvatrema chaii adults from experimental mice at week 1 after infection
Table 2.
|
Mouse no. |
No. of metacercariae infected |
No. of worms recovered from
|
|
APa)
|
MPb)
|
PPc)
|
Total (%) |
|
1 |
300 |
52 |
3 |
1 |
56 (18.7) |
|
2 |
300 |
17 |
55 |
8 |
80 (26.7) |
|
3 |
300 |
3 |
61 |
1 |
65 (21.7) |
|
4 |
300 |
88 |
12 |
1 |
101 (33.7) |
|
Total |
1,200 |
160 |
131 |
11 |
302 (25.2) |
Citations
Citations to this article as recorded by

- Parvatrema spp. (Digenea, Gymnophallidae) with parthenogenetic metacercariae: diversity, distribution and host specificity in the palaearctic
Kirill V. Galaktionov, Anna Gonchar, Daria Postanogova, Aleksei Miroliubov, Semen Yu. Bodrov
International Journal for Parasitology.2024;[Epub] CrossRef - Parvatrema duboisi (Digenea: Gymnophallidae) Life Cycle Stages in Manila Clams, Ruditapes philippinarum, from Aphae-do (Island), Shinan-gun, Korea
Bong-Kwang Jung, Taehee Chang, Hyejoo Shin, Seungwan Ryoo, Sooji Hong, Jeonggyu Lee, Hyemi Song, Jaeeun Cho, Deok-Gyu Kim, Hojong Jun, Min-Jae Kim, Eun Jeong Won, Eun-Taek Han, Eun-Hee Shin, Jong-Yil Chai
The Korean Journal of Parasitology.2021; 59(1): 83. CrossRef - Morphological and molecular characterization of larval digenean trematodes (Parvatrema: Gymnophallidae) and their pathological effects on the clam Leukoma thaca (=Protothaca thaca) (Bivalvia:Veneridae) (Molina, 1782) from northern Chile
Diana Montenegro, María Soledad Romero, María Teresa González
Parasitology International.2021; 80: 102238. CrossRef - Morphological and Molecular Confirmation of Parvatrema duboisi Metacercariae in the Manila Clam Ruditapes philippinarum from Gochang-gun, Korea
Taehee Chang, Bong-Kwang Jung, Hyejoo Shin, Sooji Hong, Jeonggyu Lee, Deok-Gyu Kim, Laddawan Patarwut, Woon-Mok Sohn, Jong-Yil Chai
The Korean Journal of Parasitology.2020; 58(1): 87. CrossRef - Prevalence and Density of Digenetic Trematode Metacercariae in Clams and Oysters from Western Coastal Regions of the Republic of Korea
Woon-Mok Sohn, Byoung-Kuk Na, Shin-Hyeong Cho, Won-Ja Lee
The Korean Journal of Parasitology.2017; 55(4): 399. CrossRef -
Enigmatic septa in shells of some Middle Jurassic
Pholadomya
(Bivalvia) from Poland
Przemysław Sztajner
Lethaia.2016; 49(3): 351. CrossRef - Two new species of Parspina Pearse, 1920 (Digenea: Cryptogonimidae) from freshwater fishes (Gymnotiformes) of the Paraná River basin in Argentina
Margarita C. Ostrowski de Núñez, Nathalia J. Arredondo, Alicia A. Gil de Pertierra
Systematic Parasitology.2011; 80(1): 67. CrossRef - Redescription of Parspina argentinensis (Szidat, 1954) (Digenea: Cryptogonimidae) from freshwater fishes (Pimelodidae) in the basins of the Paraná and La Plata Rivers, Argentina, with comments on P. bagre Pearse, 1920
Margarita C. Ostrowski de Núñez, Nathalia J. Arredondo, Irene L. Doma, Alicia A. Gil de Pertierra
Systematic Parasitology.2011; 78(1): 27. CrossRef - Discovery ofParvatrema duboisiandParvatrema homoeotecnum(Digenea: Gymnophallidae) from Migratory Birds in Korea
Ok-Sik Chung, Hye-Jung Lee, Woon-Mok Sohn, Yun-Kyu Park, Jong-Yil Chai, Min Seo
The Korean Journal of Parasitology.2010; 48(3): 271. CrossRef - Mactra veneriformis, an Intertidal Clam, as a New Second Intermediate Host for Acanthoparyphium marilae (Digenea: Echinostomatidae)
Eun-Taek Han, Jong-Yil Chai
The Korean Journal of Parasitology.2008; 46(2): 101. CrossRef