Abstract
Toxoplasma gondii tachyzoites were isolated from the blood of an ocular patient, and have been successfully passaged in the laboratory, for over a year, by peritoneal inoculation in mice. The isolated parasite was designated the Korean Isolate-1 (KI-1) and its characteristics were compared with those of the RH strain, a well-known virulent strain originating from a child who suffered from encephalitis. The morphology, pathogenicity, infectivity and cell culture characteristics of the KI-1 were similar to those of the RH strain. Both RH and KI-1 antigens were detected by an anti-T. gondii monoclonal antibody (mAb), Tg563, against the major surface protein SAG1 (30 kDa), whereas no reaction was observed against an anti-Neospora caninum mAb, 12B4. The KI-1 was confirmed as an isolate of T. gondii. A long-term laboratory maintenance and characterization of a local T. gondii isolate is reported for the first time in the Republic of Korea.
-
Key words: Toxoplasma gondii Korean Isolate-1 (KI-1) and RH strain, ocular patient, immunoblotting, culture (in vitro), passage (in mice)
INTRODUCTION
Toxoplasma gondii is an intracellular protozoan that can cause significant morbidity and mortality in man and animals (
Chai et al., 1997). In the Republic of Korea, the seroprevalence of
T. gondii was reported to be around 2-7% among the general population in the 1980s (
Choi WY et al., 1987), and about 7.7% in children younger than 10 years of age in the 1990s (
Kook et al., 1999). Several clinical toxoplasmosis cases have been reported (
Choi JS et al., 1980;
Choi WY et al., 1997;
Kim MH et al., 2000). The transmission of
T. gondii occurs either by ingestion of oocysts shed in feline feces, cysts contained in chronically infected tissues of pigs or by vertical transmission from mothers to newborns. In most adult populations,
T. gondii infection in general does not cause a serious illness, with the exception of immunocompromised patients. In congenital infections, however, this protozoan can cause abortion, blindness and mental retardation of the affected newborns.
For studies on various aspects of toxoplasmosis including diagnosis, treatment and immunization, local isolates of the parasites are needed. In patients infected with
T. gondii, the parasites can spread into the whole body via the blood stream. Therefore, many sample types can be used to isolate
T. gondii, including the bronchoalveolar lavage fluid (
Gadea et al., 1995), amniotic fluid and placenta (
Teutsch et al., 1980), saliva and tonsils (
Amendoeira and Coutinho, 1982), blood (
Hofflin and Remington, 1985) and cerebrospinal fluid (
Fu et al., 1997). The infection can also be transmitted from the brain to the eyes via the optic nerve (
Roberts and Mcleod, 1999).
In the Republic of Korea, the tachyzoites of
T. gondii were isolated from the blood of two ocular patients (
Choi et al., 1980). Unfortunately, however, neither of the two isolates was kept long enough for a long-term maintenance and characterization. We report here a successful isolation and maintenance of a Korean isolate of
T. gondii from the blood of an ocular patient. Its virulence, tissue culture characteristics and immunoblot patterns were compared with those of the RH strain, a world-wide known virulent strain, which had originated from a child suffering from encephalitis (
Høgdall et al., 2000). Our isolate was designated the Korean Isolate-1 (KI-1), and has been maintained in the laboratory, for over a year, by passages in mice.
MATERIALS AND METHODS
Brief history of the patient
A 61-year-old woman visited a local clinic with clinical complaints of visual disturbance and a retinal mass. The patient revealed a high intraocular pressure of 14 mmHg and low left eyesight of 0.2. Serological tests revealed typical patterns of an acute infection with increasing levels of IgM, 32.6 IU/ml (reference value; 0-6 IU/ml), and IgG, 5.14 IU/ml (reference value; 0-0.8 IU/ml), as measured by the enzyme immunoassay kits (Beckman Coulter, Inc., Chaska, Minnesota, U.S.A.). The antibody titer against T. gondii was 1 : 64 by the latex agglutination test (Eiken, Tokyo, Japan). The differential counts of WBC were as follows: neutrophils 27.4%, lymphocytes 58.1%, monocytes 11.7% and eosinophils 2.8%. After treatment with sulfadiazine and pyrimidine, the left eyesight of the patient increased to 0.3 and the intraocular pressure decreased to 12 mmHg. The lymphocytes and monocytes returned to their normal levels.
Isolation and maintenance of T. gondii (the KI-1)
The peripheral blood of the patient was drawn using a syringe, and transferred into a test tube. The blood was left to coagulate at room temperature, and the serum was taken out. The blood clot, with a small amount of serum, was frozen to -70℃, and thawed just before use. After thawing, the blood clot was minced with the tip of a pipette, and injected intraperitoneally into 3 young (6-week-old) female BALB/c mice. Seven days later, the mice were sacrificed and the exudates were drawn from the peritoneal cavity. The fluid was centrifuged at 2,500 rpm for 10 min at room temperature and the sediment was again inoculated intraperitoneally into 3 young mice. Several times of similar mouse passages were repeated. Each time when the peritoneal fluid was obtained, the sediment was smeared to detect T. gondii tachyzoites using a light microscope (LM) after Giemsa staining.
After identifying the presence of tachyzoites, the peritoneal fluid of the mice was inoculated into new young mice, and 3 days later the mice were sacrificed and the peritoneal fluids collected. Using this procedure the KI-1 isolate has been continuously maintained in our laboratory.
Transmission electron microscopy of the KI-1 tachyzoites
The KI-1 tachyzoites, obtained from the peritoneal fluid of BALB/c mice, were processed for transmission electron microscopy (TEM), following the procedure reported previously (
Kook et al., 1995;
Chai et al., 1997). Briefly, the tachyzoites were washed with cacodylate buffer (pH 7.4), and fixed with 2.5% glutaraldehyde at 4℃ for 12 hrs. They were post-fixed with 1% osmium tetroxide for 2 hrs, and then dehydrated in a graded series of ethanol. They were embedded in epon, and semithin and ultrathin sections were prepared. The ultrathin sections were stained with uranyl acetate and lead citrate, and observed using a TEM (1200 EX-II, JEOL, Japan) at 80 KV.
The Sarcoma 180 cell line was purchased from the Korea Cell Line Bank (Seoul, Korea). The cells were grown in RPMI 1640 medium (Gibco BRL, Grand Island, NewYork, U.S.A.) supplemented with 10% fetal bovine serum (Gibco BRL) at 37℃ in an atmosphere of 95% air and 5% CO2. The cells were observed daily using an inverted LM.
Two culture flasks containing the Sarcoma 180 cells were each infected with 1 × 10
6 tachyzoites of the KI-1. A host cell-parasite ratio of 3 : 1 was used, as this ratio has been shown to yield a large number of intracellular parasites (
Couatarmanach et al., 1991). The cell suspensions were collected from the flasks daily, by centrifugation at 1,500 rpm for 5 min from day 1 to day 6 post-infection (PI). The numbers of tachyzoites and cells in the decanted supernatant were counted in a hemocytometer and their viability determined by the trypan blue exclusion test. To avoid cell damages due to acidification of the medium, the RPMI 1640 medium was changed everyday.
Six-week-old, specific pathogen-free, BALB/c mice (females) were purchased from the Laboratory Animal Center in Seoul National University (Seoul, Korea), and intraperitoneally inoculated with variable doses (103, 104, 105, 106, 107 and 108) of T. gondii KI-1 and RH tachyzoites. The survival times of the mice were recorded from infection until day 8 PI.
Preparation of T. gondii and Neospora caninum antigens
The RH and KI-1 tachyzoites of
T. gondii, harvested from the peritoneal exudates of experimentally infected mice, were purified using 40% Percoll (Pharmacia Biotech, Uppsala, Sweden) in phosphate buffered solution (PBS) (pH = 7.4) (
Choi WY et al., 1992). The purified tachyzoites were disrupted by five cycles of freezing to -70℃ and thawing at room temperature. The homogenate was centrifuged at 12,000 rpm at 4℃ for 30 min and the supernatant used as the antigen.
The KBA2 isolate of
N. caninum (
Kim JH et al., 2000) was obtained from the Department of Pathology, Seoul National University College of Veterinary Sciences, and has been maintained in our laboratory by a previously described procedure (
Kim JH et al., 2000). The procedure for preparation of the
N. caninum antigen was the same as that for preparation of the
T. gondii antigen.
The protein bands of the RH, KI-1 and
N. caninum antigens were separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) on 10% gels and transferred onto polyvinylidene fluoride nitrocellulose membranes (Millipore Corporation, Bedford, Massachusetts, U.S.A.) (
Towbin et al., 1979). The membranes were incubated at room temperature for 2 hrs with an anti-
T. gondii monoclonal antibody (mAb), Tg563, at a 1:100 dilution (Catholic University of Korea, Seoul, Korea) and an anti-
N. caninum mAb, 12B4, at a 1:100 dilution (Catholic University of Korea, Seoul, Korea). Horseradish peroxidase-conjugated goat anti-mouse serum (Cappel Company, St. Louis, Missouri, U.S.A.), at a 1:500 dilution, was used as the antibody. The blots were developed with 4-chloro-1-naphthol containing 0.03% H
2O
2 in phosphate buffer (0.01M, pH 7.4).
RESULTS
LM and TEM morphology of the KI-1 tachyzoites
Seven days after the second passage of the patient's blood clot into young BALB/c mice, the mice were sacrificed, and several drops of the peritoneal exudate were smeared on a glass slide and examined for
Toxoplasma using a LM after Giemsa staining.
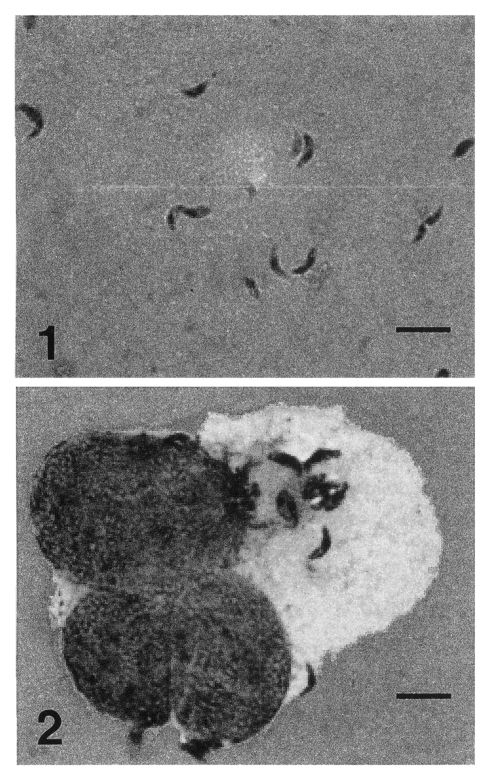
Toxoplasma tachyzoite-like organisms were found in considerable numbers in 1 of the 3 mice. They were pyriform or crescent shaped (
Figs. 1-2), measuring 4 to 6 µm long and 2 to 3 µm wide, each having an apparent nucleus. Their morphology was similar to that of the RH tachyzoites of
T. gondii.
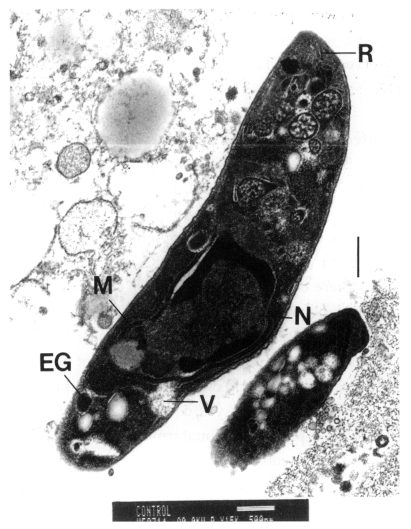
The TEM morphology of the KI-1 tachyzoites (
Fig. 3) was consistent with that of the RH tachyzoites of
T. gondii (
Kook et al., 1995;
Chai et al., 1997). The pellicle of the parasite was composed of two membranes forming sharp cell surfaces. The number of rhoptries was 6 to 8 in each tachyzoite. The nucleus had dense chromatins in the center as well as along the nuclear wall, and thus it was easily differentiated from the cytoplasm. Electron dense granules, ribosomes, vacuoles, and mitochondria were also seen (
Fig. 3).
The Sarcoma 180 cells grew in suspension cultures as non-adherent polymorphic cells. The cell line was successfully infected by the KI-1 and intracellular multiplication of the parasites occurred. The KI-1 tachyzoites in the Sarcoma 180 cells (
Fig. 2) presented a similar morphology to that observed in mice in vivo (
Fig. 1). When 1 × 10
6 tachyzoites were seeded into the Sarcoma 180 cell suspension, containing 3 × 10
6 cells in RPMI medium, there was a 256-fold increase in the number of tachyzoites at day 5 PI; the number of tachyzoites and Sarcoma 180 cells being 2.56 × 10
8 and 1.23 × 10
5, respectively.
The virulence of the KI-1 tachyzoites was estimated by observing the survival of the mice after intraperitoneal inoculation, and compared with that of the RH tachyzoites (
Table 1). At the inoculum dose of 10
8 tachyzoites per mouse, only two of five (40.0%) and three of five (60.0%) mice, for the RH and KI-1 groups, respectively, survived to day 2 PI, and no mice survived longer than 3 days from either group (
Table 1). At the dose of 10
7 tachyzoites per mouse, mice in both groups survived equally, with the survival rate of 60.0% at day 3 PI. At the dose of 10
6 tachyzoites per mouse, however, the survival rate on day 4 PI was different between the two groups, 40.0% and 80.0% for the RH and KI-1 groups, respectively. At the dose of 10
5 tachyzoites per mouse, the survival rate on day 5 PI was 40.0% and 60.0%, and that on day 6 PI was 0% and 20.0%, for the RH and KI-1 groups, respectively. The survival of mice was similar between the two groups at the inoculum doses of 10
4 and 10
3 tachyzoites per mouse. These results showed that the KI-1 is an isolate with strong virulence, comparable to that of the RH strain.
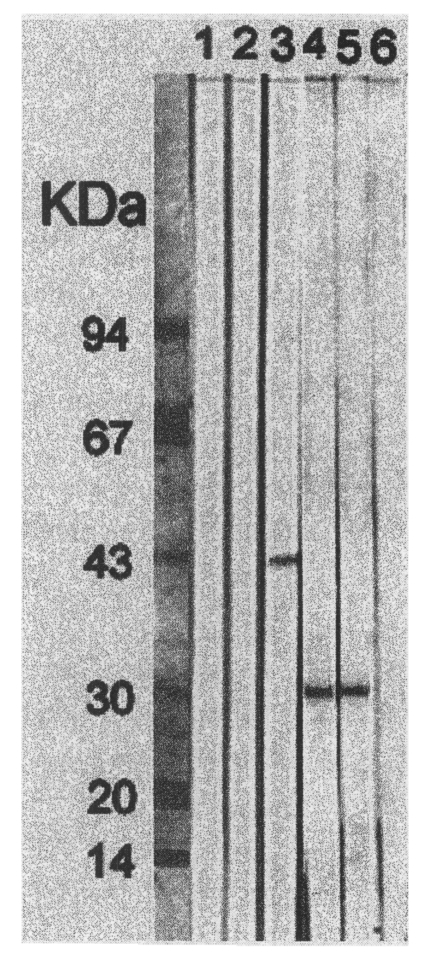
Immunoblotting of the RH and KI-1 antigens showed that both antigens reacted to the mAb (Tg 563) against the
T. gondii SAG1 antigen (30 kDa), but not to the mAb (12B4) against the
N. caninum antigen (
Fig. 4). The mAb, 12B4, reacted only to the
N. caninum antigen (
Fig. 4).
DISCUSSION
Relatively lower prevalences and antibody titers of
T. gondii have been reported among Koreans compared with U.S. and Europian populations (
Choi et al., 1992;
Kook et al., 1999). This might be related to the food-eating habits (rarely consume raw or improperly cooked pork) and pet-loving fashions (prefer puppies to kittens) of the Korean people. In addition, clinicians who are working in the field of infectious diseases in the Republic of Korea have paid little attention to toxoplasmosis. Moreover, the sensitivity of serological tests may be low;
T. gondii may be present in the saliva of patients in early stages of infection, when the infection cannot be detected by the serological tests (
Amendoeira and Coutinho, 1982). Isolation of the
T. gondii parasite is necessary in such patients, whether their serological tests are positive or negative.
In the Republic of Korea, the organism responsible for human toxoplasmosis was isolated from 4 patients, i.e., 2 congenital (
Chung et al., 1980) and 2 chorioretinitis patients (
Choi et al., 1980). Unfortunately, however, the properties of the isolates were not characterized, and the isolates were not maintained in the laboratory.
The present study describes, for the first time in the Republic of Korea, a successful isolation and characterization of
T. gondii from a patient, with long-term (over a year) laboratory maintenance of the isolate (KI-1) by mouse inoculation. The morphology, antigenicity and virulence of the KI-1 tachyzoites were similar to those of the RH strain (
Kook et al., 1995;
Chai et al., 1997). The KI-1 survived and actively multiplied in the Sarcoma 180 cells; a 256-fold increase in the number of tachyzoites was observed at day 5 PI. Immunoblotting revealed that both the RH and KI-1 antigens reacted specifically to an anti-
T. gondii mAb, but not to an anti-
N. caninum mAb. Based on these results, the KI-1 was confirmed to be an isolate of
T. gondii.
Since
T. gondii was first described in a rodent, numerous strains, or isolates, have been obtained from humans. For example,
T. gondii was isolated from the saliva and tonsils of a 3-year-old child and maintained by mouse inoculation (
Amendoeira and Coutinho, 1982). Tachyzoites were obtained from the blood of an acquired immunodeficiency syndrome (AIDS) patient, and maintained by tissue cultures (
Hofflin and Remington, 1985). Three isolates were obtained from the cerebrospinal fluid of humans and have been maintained by mouse inoculation at the Zhejiang Academy of Medical Sciences, China (
Fu et al., 1997).
The strains of
T. gondii were clustered into 2 major virulence groups, virulent (including the RH) and avirulent (including the ME49), with the virulent strains comprising of only a single clonal lineage (
Sibley and Boothroyd, 1992). More recently, however, 3 clonal lineages were identified among
T. gondii using a polymerase chain reaction-based restriction fragment length polymorphism (PCR-RFLP) analysis; type I, II and III (
Howe and Sibley, 1995). The types I and II were virulent (including the RH) and avirulent strains (including the PLK), respectively, and the type III consisted of avirulent and less virulent isolates (
Howe and Sibley, 1995). However, evidence was shown that not all the virulent isolates are closely related. For example, the strain MAS, highly pathogenic to mice, was consistent with the type I in only 4 of 6 allelic loci studied; different patterns were observed in the remaining 2 loci, i.e., L328 and 62 (
Howe and Sibley, 1995). The type I strains were most frequently seen in congenital toxoplasmosis of humans, whereas the type II predominated among AIDS patients; the type III was most common in animal infections (
Howe and Sibley, 1995). It is interesting to note that the KI-1 is a strongly virulent isolate, and seems to belong to the type I. However, studies on the DNA patterns of the KI-1 are needed.
The sequence polymorphism has also been studied in a number of DNA loci of different
T. gondii strains, including the genes for the heat-shock protein 70 (
Lyons and Johnson, 1998), a dense granule antigen GRA6 (
Fazaeli et al., 2000a) and the intergenic spacer (
Fazaeli et al., 2000b). The sequence variants of
T. gondii for those genes were used as markers for distinguishing
T. gondii isolates from different animals and geographical localities (
Høgdall et al., 2000). Therefore, analyses of the sequence heterogeneity of the KI-1 genes should be performed. Research on the development of diagnostic kits and vaccines using the KI-1 is also needed.
Notes
-
This work was supported by grant No. R01-2002-000-00422-0 from the Basic Research Program of the Korea Science and Engineering Foundation.
ACKNOWLEDGMENTS
We would like to thank Prof. Dae-Yong Kim, Department of Pathology, Seoul National University College of Veterinary Medicine, Seoul, Korea, for providing us with the Neospora caninum KBA2 strain.
References
- 1. Amendoeira MRR, Coutinho SG. Isolation ofToxoplasma gondii from the saliva and tonsils of a three-year-old child. J Infect Dis 1982;145:587.
- 2. Chai JY, Kook J, Guk SM, Chang YP, Yun CK. Experimental infection of murine splenic lymphocytes and granulocytes with Toxoplasma gondii RH tachyzoites. Korean J Parasitol 1997;35:79-85.
- 3. Choi JS, Choi CS, Soh CT. Isolation of Toxoplasma gondii from congenital and acquired chorioretinitis cases. Yonsei Rep Trop Med 1980;11:39-42.
- 4. Choi WY, Nam HW, Kwak NH, et al. Foodborne outbreaks of human toxoplasmosis. J Infect Dis 1997;175:1280-1282.
- 5. Choi WY, Nam HW, Youn JH, et al. Detection of antibodies in serum and cerebrospinal fluid to Toxoplasma gondii by indirect latex agglutination test and enzyme-linked immunosorbent assay. Korean J Parasitol 1992;30:83-90.
- 6. Choi WY, Yoo JE, Nam HW, et al. Toxoplasma antibody titers by indirect latex agglutination tests in patients of Kangnam St. Mary's Hospital Cheju Medical Center. Korean J Parasitol 1987;25:13-23.
- 7. Chung KS, Kung RN, Chung KS, Kim PK, Yun DJ, Soh CT. Congenital toxoplasmosis. Yonsei Med J 1980;21:62-74.
- 8. Couatarmanach A, Andre P, Le Minous D, Martin L, Robert R, Deunff J. In vitro culture and cloning of Toxoplasma gondii in a newly established cell line derived from TG180. Int J Parasitol 1991;21:129-132.
- 9. Fazaeli A, Carter PE, Darde ML, Pennington TH. Molecular typing of Toxoplasma gondii strains by GRA6 gene sequence analysis. Int J Parasitol 2000a;30:637-642.
- 10. Fazaeli A, Carter PE, Pennington TH. Intergenic spacer (IGS) polymorphism: a new genetic marker for differentiation of Toxoplasma gondii strains and Neospora caninum. J Parasitol 2000b;86:716-723.
- 11. Fu CE, Yang JS, Chen CH, et al. Study on quick diagnostic method of Toxoplasma gondii of central nervous system. Chin J Zoon 1997;13:31-33. (in Chinese).
- 12. Gadea I, Cuenca M, Benito N, Pereda JM, Soriano F. Bronchoalveolar lavage for the diagnosis of disseminated toxoplasmosis in AIDS patients. Diagn Microbiol Infect Dis 1995;22:339-341.
- 13. Hofflin JM, Remington JS. Tissue culture isolation of Toxoplasma from the blood of a patient with AIDS. Arch Intern Med 1985;145:925-926.
- 14. Høgdall E, Vuust J, Lind P, Petersen E. Characterization of Toxoplasma gondii isolates using polymerase chain reaction (PCR) and restriction fragment length polymorphism (RFLP) of the non-coding Toxoplasma gondii (TGR)-gene sequences. Int J Parasitol 2000;30:853-858.
- 15. Howe DK, Sibley LD. Toxoplasma gondii comprises three clonal lineages: Correlation of parasite genotype with human diseases. J Infect Dis 1995;172:1561-1566.
- 16. Kim JH, Sohn HJ, Hwang WS, et al. In vitro isolation and characterization of bovine Neospora caninum in Korea. Vet Parasitol 2000;90:147-154.
- 17. Kim MH, Choi YK, Park YK, Nam HW. A toxoplasmic uveitis case of a 60-year-old male in Korea. Korean J Parasitol 2000;38:29-31.
- 18. Kook J, Kim YJ, Seo M, Chai JY. TEM ultrastructure of gamma-irradiated Toxoplasma gondii RH tachyzoites. Seoul J Med 1995;36:159-165.
- 19. Kook J, Lee HJ, Kim BI, et al. Toxoplasma gondii antibody titers in sera of children admitted to the Seoul National University Children's Hospital. Korean J Parasitol 1999;37:27-32.
- 20. Lyons RE, Johnson AM. Gene sequence and transcription differences in 70kDa heat shock protein correlates with murine virulence of Toxoplasma gondii. Int J Parasitol 1998;28:1041-1051.
- 21. Roberts F, Mcleod R. Pathogenesis of toxoplasmic retinochoroiditis. Parasitol Today 1999;15:51-57.
- 22. Sibley LD, Boothroyd JC. Virulent strains of Toxoplasma gondii comprise a single clonal lineage. Nature 1992;359:82-85.
- 23. Teutsch SM, Sulzer AJ, Ramsey JE Jr, Murray WA Jr, Juranel DD. Toxoplasma gondii isolated from amniotic fluid. Obstet Gynecol 1980;55(3 suppl):2S-4S.
- 24. Towbin M, Staebelin T, Gordon J. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheet: procedure and some applications. Proc Natl Acad Sci 1979;76:4350-4354.
Figs. 1-2The Korean Isolate-1 (KI-1) tachyzoites of Toxoplasma gondii isolated from an ocular patient. Giemsa stained. Scale bar = 12 µm. Fig. 2. The KI-1 tachyzoites of T. gondii in the Sarcoma 180 cell line cultured in vitro. Giemsa stained. Scale bar = 10 µm.

Fig. 3TEM morphology of a KI-1 tachyzoite. The nucleus has dense chromatins in the center and along the nuclear wall, and are easily differentiated from the cytoplasm. Characteristic organelles such as the nucleus (N), rhoptries (R), electron dense granules (EG), vacuoles (V) and mitochondria (M) are seen. Scale bar = 0.5 µm.

Fig. 4Immunoblot of the RH and KI-1 of T. gondii and Neospora caninum KBA2 antigens, with the anti-Toxoplasma monoclonal antibody (mAb) Tg563 and anti-Neospora mAb 12B4. The RH, KI-1 and KBA2 antigens were reacted with the 12B4 (Lanes 1, 2 and 3), and the RH, KI-1 and KBA2 antigens were reacted with the Tg563 (Lanes 4, 5 and 6).

Table 1.Survival rates of mice infected with variable doses of Toxoplasma gondii RH or KI-1 tachyzoites
Table 1.
|
Inoculum dose of tachyzoites |
T. gondii strain or isolate |
Survival rates by day after tachyzoite inoculation (%)
|
|
2 |
3 |
4 |
5 |
6 |
7 |
8 |
|
108
|
RH |
40a)
|
0 |
0 |
0 |
0 |
0 |
0 |
|
KI-1 |
60 |
0 |
0 |
0 |
0 |
0 |
0 |
|
107
|
RH |
100 |
60 |
0 |
0 |
0 |
0 |
0 |
|
KI-1 |
100 |
60 |
0 |
0 |
0 |
0 |
0 |
|
106
|
RH |
100 |
100 |
40 |
0 |
0 |
0 |
0 |
|
KI-1 |
100 |
100 |
80 |
0 |
0 |
0 |
0 |
|
105
|
RH |
100 |
100 |
100 |
40 |
0 |
0 |
0 |
|
KI-1 |
100 |
100 |
100 |
60 |
20 |
0 |
0 |
|
104
|
RH |
100 |
100 |
100 |
60 |
20 |
0 |
0 |
|
KI-1 |
100 |
100 |
100 |
60 |
40 |
0 |
0 |
|
103
|
RH |
100 |
100 |
100 |
100 |
40 |
40 |
0 |
|
KI-1 |
100 |
100 |
100 |
60 |
40 |
40 |
0 |