Abstract
Antibody responses in serum and cerebrospinal fluid (CSF) samples from patients with active and chronic paragonimiasis and in sera from patients on whom follow-up studies were done after praziquantel treatment were analyzed using antigens of Paragonimus westermani prepared from eggs, metacercariae, juveniles of 4- and 7-week old, adult worms and recombinant protein of 28 kDa cruzipain-like cysteine protease (rPw28CCP). The patient sera/CSFs of active and chronic paragonimiasis revealed strong antibody reactions against the crude extracts of 4- and 7-week old juveniles as well as against those from egg and adult. rPw28CCP also showed specific reaction to the sera with active paragonimiasis. After the treatment, levels of specific antibodies in the sera gradually decreased to negative range in most patients. In some cases with persisting high antibody levels, however, the reactions at 27 kDa egg protein were sustained throughout the observation period of 34 months. The reactions at 35 and 32 kDa in adult extract and rPw28CCP disappeared rapidly after the treatment. Persistent antibody reactions even after successful treatment are provoked by continuous antigenic challenge from eggs which were not resolved by treatment.
-
Key words: Paragonimus westermani, paragonimiasis, chemotherapy, follow-up, epitope change
INTRODUCTION
Paragonimiasis, which is caused by the lung fluke,
Paragonimus westermani, and other species is still endemic worldwide and shows high incidence in East and Southeast Asia (
Cho et al., 1997). When infective metacercariae are ingested, they penetrate the duodenum and the peritoneal cavity, and migrate to the lungs. Some of infected worms migrate aberrantly and elicit ectopic paragonimiasis in the abdominal, subcutaneous or brain tissues (
Blair et al., 1999). Nowadays, the significance of differential diagnosis of paragonimiasis from pulmonary tuberculosis has increasingly been recognized (
WHO, 1995).
The serological diagnosis of paragonimiasis has been directed to detect
Paragonimus-specific antibodies in the patient sera (
Cho et al., 1981;
Ikeda, 1998a;
Kong et al., 1998). The antibody tests have been reported to be highly sensitive compared to egg detection, also being able to diagnose ectopic infections. In serological diagnosis of paragonimiasis, crude extracts of the adult worms have widely been used (
Cho et al., 1981;
1989;
Slemenda et al., 1989;
Kong et al., 1998). A partially purified or a recombinant cysteine protease has recently appeared to have high reliability in diagnosing active paragonimiasis (
Ikeda et al., 1996;
Ikeda, 1998a,
1998b;
Yun et al., 2000). The crude extracts of adult worms can diagnose wide clinical spectrum of the paragonimiasis. However, as a specific diagnostic antigen, the crude extract has not well been studied, especially regarding its cross-reactivity with other parasitic diseases.
As for the serological follow-up study to identify the cure criteria for paragonimiasis, observing that the antibody reactions by complement fixation test or by enzyme-linked immunosorbent assay (ELISA) has reported the return of specific antibody levels to negative ranges within 7 months after specific treatment (
Yokogawa, 1965;
Knobloch et al., 1984). Cho et al. (
1989), also, showed that the antibody reactions studied by ELISA converted to negative range in a variable period from 4 to 28 months with or without temporary elevations in 2-6 months after praziquantel treatment. We have experienced some cases, however, in which the antibody levels did not return to negative ranges even after the presumably successful treatment although the overall antibody levels gradually decreased. Such examples suggested that the existing covert paragonimiasis lesions might provoke the persistent antibody responses by continuously releasing small dose of antigenic materials. The first candidate of antigenic source is the eggs because they do not resolve by drug treatment.
In the present study, we observed whether the eggs of P. westermani are main causes of persisting antibody levels in some cured cases of paragonimiasis. For the purpose, several different antigens prepared from eggs, metacercariae, juveniles, adult and recombinant cruzipain-like cysteine protease of adult P. westermani (rPw28CCP) were used to examine their capability of antibody recognition in a variety of human clinical paragonimiasis including follow-up cases.
MATERIALS AND METHODS
Preparation of different antigens of Paragonimus westermani
The crude extracts of different developmental stages of
P. westermani including newly excysted metacercariae, 4- and 7-week old juveniles, 16-week old adults, and eggs were prepared as described previously (
Chung et al., 1995,
1997;
Kang et al., 1995;
Kong et al., 1998). In brief, the metacercariae were collected from naturally infected crayfish,
Cambaroides similis. They were excysted using 5 mM dithiothreitol (DTT, Sigma) at 37℃ in Tris-HCl buffer (0.1 M, pH 8.0). Four-week old worms were harvested from the peritoneal, thoracic and pleural cavities of infected cats. The 7-week old juvenile and adult worms were collected from the lungs of infected dogs. Eggs were obtained by incubating the adult worms in physiological saline at 37℃ in CO
2 incubator overnight. They were homogenized with a teflon-pestle tissue homogenizer in physiological saline, after which the homogenates were centrifuged twice at 700 g for 5 min followed by 20,000 g for 1 hr. Resulting supernatants were used as crude extracts of eggs and worms of different age. All procedures for protein extraction were carried out at 4℃ unless otherwise specified.
A recombinant protein of the 28 kDa cruzipain-like cysteine protease of adult
P. westermani (rPw28CCP) was expressed bacterially as described elsewhere (
Yun et al., 2000). In brief, a 642-bp mature protein domain was amplified by polymerase chain reaction with gene specific primers 5'-AGCT
CATATGGCCCCGGCAAGTGTTGACTG-3' and 5'-GAAGT
CTCGAGTTAGTGAATGATGGCG G-3', incorporating
NdeI and
XhoI site (underlined sequence each) to aid the purification. The recombinant plasmid was inserted to pET28a (+)
E. coli expression vector. The fidelity of expression was confirmed by DNA sequencing. Upon induction with 0.1 mM isopropylthiol-β-D-galactoside, the cells were harvested and lysed using TE buffer containing 10 µg/ml of lysozyme. The recombinant protein expressed at a high level as inclusion bodies was dissolved by 6 M urea. The protein was purified by His-Bind metal chelation resin (Novagen, Madison, WI, USA). Urea was removed by stepwise dialysis in the presence of 2 mM DTT. The protein migrated at 27 kDa as described elsewhere (
Yun et al., 2000).
All the antigenic materials prepared was stored at -70℃ or in liquid nitrogen until use.
Serum and cerebrospinal fluid (CSF) samples used
Sera from cases with active paragonimiasis:
A total of 16 sera from paragonimiasis patients who complained of cough, hemoptysis, chest/pleuritic pain and/or sputum were selected from our sera bank. They were diagnosed by positive antibody reaction to enzyme-linked immunosorbent assay (ELISA) together with typical findings of high resolution chest CT (
Im et al., 1992,
1993), or by egg detection.
Sera and CSFs of cerebral paragonimiasis patients:
The sera and CSFs from cases with chronic cerebral paragonimiasis patients who showed typical neuroimaging findings in their brain CT/MR (
Cha et al., 1994;
Nomura et al., 1999) together with positive antibody reactions in ELISA performed with their sera and CSFs were subjected to the test. Their chief complaints included headache, seizure, hemiparesis, focal neurological deficit and/or mental impairment. From 22 cases with chronic calcified cerebral paragonimiasis, 22 serum samples and 13 CSFs were collected and used in this study. The sera and CSFs were stored at -70℃ until use.
Patient sera from follow-up examinations:
A total of 98 sera from 19 patients, who had been followed-up after praziquantel treatment, was included to the test to observe the changing patterns of the specific antibodies in their sera. Follow-up period varied from 5 to 34 months.
Enzyme linked immunosorbent assay (ELISA)
The specific IgG antibody levels in sera/CSFs from the patients were detected by micro-ELISA as described elsewhere (
Cho et al., 1981). In brief, each antigen (200 µl, protein contents 2.5 µg/ml) was coated overnight at 4℃ in microtiter plate (Costar, Cambridge, CA, USA). The sera diluted at 1:100 and neat CSFs were also incubated for 2 hr at 37℃. Peroxidase conjugated anti-human IgG (heavy- and light-chain specific, Cappel, West Chester, PA, USA) was diluted at 1:1,000 and incubated further for 2 hr at 37℃. Color reaction was developed by o-phenylene diamine chromogen (Sigma). The absorbance was read at 490 nm. Absorbance (abs) at 0.25 was used as the positive criteria in both serum and CSF (
Cho et al., 1981).
Patient sera (20 µl) were reacted with preactivated Pansorbin (20 µl, Calbiochem, San Diego, CA, USA) for 2 hr at 4℃. The adult extract (30 µl) was added and further incubated for 2 hr at 4℃. The immune complex was washed with PBS/T for 5 times by pelleting at 15,000 g for 5 min prior to subject to SDS-polycrylamide gel electrophoresis (PAGE). The proteins resolved by SDS-PAGE were further processed with immunoblot (
Yang et al., 1998). The pooled serum of 10 cases with active paragonimiasis or monoclonal antibody (mAb) specific to egg protein, diluted at 1:200 and 1:100, respectively, was incubated overnight as primary antibody. Peroxidase-conjugated anti-human IgG (heavy- and light-chain specific, Cappel) or peroxidase-conjugated anti-mouse IgG (whole molecule, Cappel) was reacted for over 4 hr at 1:1,000 dilution. Blots were developed by 4-chloro-1-naphthol chromogen (Sigma). To characterize the mAb generated, immunoblot was carried out essentially as described above.
A total of 5 BALB/c mice was immunized 3 times with the adult worm extract mixed with Freund adjuvant (Sigma) as previously described (
Kang et al., 1991). Murine splenocytes were hybridized with SP2/0 plasmacytoma cell in the presence of 50% polyethylene glycol (E. Merck, Germany). Fused cells were cultured in HAT and HT media. Antibody secreting colonies were dispensed by limiting dilution. A clone that produced specific antibody against egg protein was selected and expanded in 15 ml culture flask (Corning, Cambridge, MA, USA). IgG fractions were isolated by protein A-Sepharose 4B affinity chromatography (Pharmacia-LKB, Uppsala, Sweden). Isotype of the mAb was determined by sandwich-ELISA using Boeringer Manheim isotyping kit with goat anti-mouse IgG1, IgG2a, IgG2b, IgG3, IgM and IgA as recommended by manufacturer's instruction (Boeringer Manheim, Indianapolis, IN, USA). The purified mAb was stored at - 70℃ until use.
RESULT
Characterization of the monoclonal antibody (mAb)
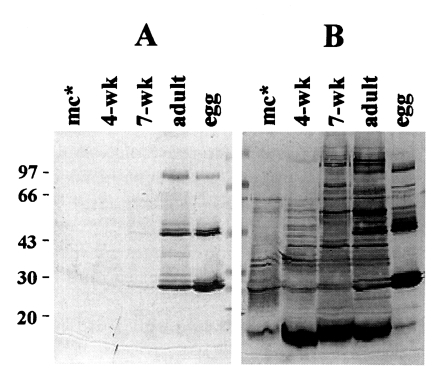
Fig. 1 shows the immunoblot analysis of mAb raised against egg protein. The mAb specifically recognized 94, 46, 28 and 27 kDa epitopes in extracts of both egg and adult worm while it did not react with extracts from metacercaria and 4-wk old juveniles (
Fig. 1A). The extract from 7-week old shows a faint band at 27 kDa, suggesting that the immature eggs have started to be produced. In contrast, when the same blot was probed with anti-serum from rabbit immunized with the adult worm extract, numerous bands reacted with polyclonal antibody (
Fig. 1B). The mAb raised in this study was found to be identical with that by Kang et al. (
1991). This also corresponded well with previous results in which the purified egg proteins showed the same protein bands in reducing SDS-PAGE analysis (
Kang et al., 1995). Isotyping of the mAb by ELISA revealed abs of 0.72 to IgG, 1.07 to IgG1, 0.26 and 0.12 to κ- and λ-chain of IgG1, respectively. Other subclasses such as IgA, IgM, IgG2a and IgG2b showed negligible abs. The mAb generated was an IgG1 type.
Specific antibody levels in active pulmonary and chronic cerebral paragonimiasis
Specific IgG antibody levels in active paragonimiasis against different antigenic preparations measured by micro-ELISA are demonstrated in
Table 1. The sera from the cases of active paragonimiasis revealed high levels of antibodies against 4- and 7-week old juveniles and adult extracts which comparable to each other, while exhibiting low levels of specific antibodies against metacercarial extract. Similar result was observed in the cases of chronic cerebral paragonimiasis. The highest levels of the antibodies in both sera and CSFs were observed using the 4-week old juvenile extract as an antigen (0.69±0.37 in sera and 0.47±0.27 in CSF, respectively). Adult and egg extracts showed high antibody levels as well (0.49±0.32 in sera and 0.43±0.32 in CSF, respectively).
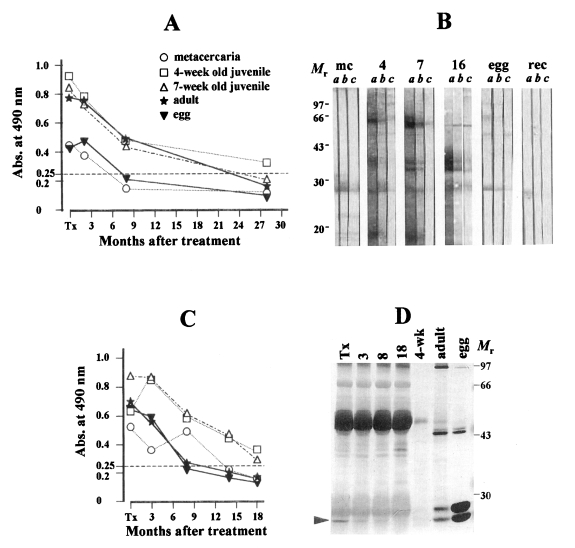
Representative changing patterns of the anti-
Paragonimus specific antibody levels analyzed by ELISA with the sera collected serially after the treatment with praziquantel against differently prepared antigens were shown: One with rapid decline (
Fig. 2A, 2C) and another with persistent antibody reactions in prolonged period (
Fig. 3A, 3C).
Figs. 2A and 2C exhibited the typical pattern of rapid decline of the antibody levels with or without temporary elevation. Out of 19 followed-up patients, 11 cases (57.8%) exhibited this pattern of decline. The specific antibody levels in the sera of this group decreased gradually according to time lapse regardless of any antigenic preparations tested in this study. In these cases, antibody responses against 4-week old worm were the highest when compared to other antigenic preparations, while those against egg and metacercarial extracts were low.
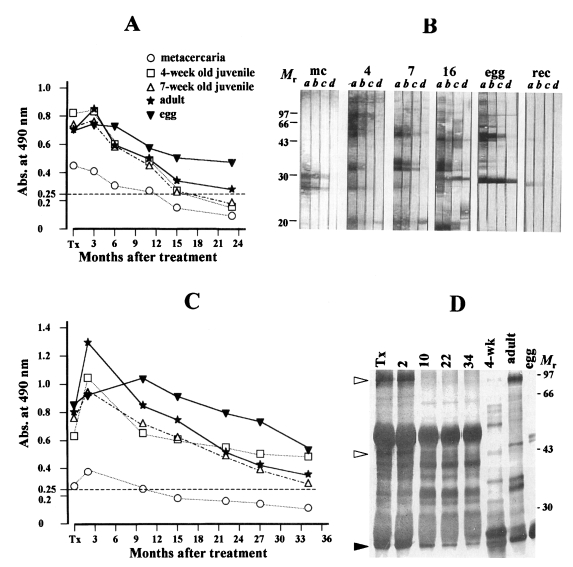
Fig. 3A and 3C illustrated another declining pattern of antibody levels in which sustained positive antibody reactions were observed for a prolonged period. Out of 19 followed-up patients, eight patients (42.2%) exhibited this pattern. The specific antibody levels in the sera of these cases were the highest against the egg extract when compared to those against the extracts of adults, 4-week old juveniles, 7-week old juveniles and metacercariae (in order of sequence). However, they also showed the declining patterns with or without temporary elevation throughout the observation period of 34 months after treatment.
Epitope recognition in the sera of follow-up monitoring after treatment
As previously reported, most sera from patients with active pulmonary paragonimiasis exhibited specific immunoreactive bands at 35 and 32 kDa in the 7-week and adult extracts (
Maleewong et al., 1997;
Kong et al., 1998) as they did in early experimental cat paragonimiasis (
Kong et al., 1997). In addition, 94, 46, 28 and 27 kDa in the adult extract exhibited positive antibody reactions (
Kong et al., 1998).
To observe the specific epitopes which were responsible for sustained antibody responses, immunoblot or immunoprecipitation analysis was performed using sera collected serially after treatment.
Fig. 2B showed the immunoblot analysis in which the follow-up sera from a patient were reacted with differentially prepared antigens of
P. westermani. The sera used in this experiment were the same sera used in
Fig. 2A. At the time of diagnosis (lane a of each panel), the patient serum reacted weakly with 28 and 27 kDa proteins of the metacercaria. The proteins at 66 kDa of 4-week old worm, at 66, 35 and 32 kDa in 7-week old worm and at 35 and 32 kDa in adult extracts showed strong antibody reactions. Reactions at 27 kDa protein of both egg extracts and recombinant protein were evident, but weak. As the time passed, all reactive protein bands disappeared without any significant changes in epitope recognition in follow-up sera of 8 and 28 months post-treatment. In
Fig. 2D, sera from a patient (same patient sera shown in
Fig. 2C) who was followed-up for 18 months after treatment were immunoprecipitated with adult worm extract and analyzed by immunoblot using the mAb against egg protein. Reactions at 27 kDa egg protein disappeared after 8 months post-treatment.
Fig. 3C exhibited another pattern of declining antibody levels as assessed by immunoblot. The sera used in this experiment were the same sera used in
Fig. 3A. Reactions at 35 and 32 kDa in both 7-week old and adult worms faded out shortly after treatment in 15 months (lanes a-d, panels 7 and 16). Antibody recognition by the rPw28CCP also diminished promptly within 11 months post-treatment (panel rec). On the while, antibody reactions against 27 kDa of the egg extract persisted throughout the observation period of 23 months while those against 94 and 46 kDa disappeared in 10 months post-treatment (lanes a-d, panels 16 and egg). When the follow-up sera from other patients were subjected to immunoprecipitation with the adult extract and probed with a pooled serum of 10 cases of active pulmonary paragonimiasis, specific epitopes at 94, 46 and 27 kDa were recognized (
Fig. 3D). As absorbance decreased, immunoreactions against 94 and 46 kDa proteins became fainter and disappeared on 10 months after treatment; however, reactions at 27 kDa band persisted throughout the follow-up period of 34 months.
DISCUSSION
One of the critical aspects of the antibody test lies in its antigenic preparation because the antibody formation in mature B cells is induced by a variety of antigenic challenges released by the worm (
Garraud et al., 1997). Therefore, composition and antigenicity of soluble proteins in the diagnostic antigen may critically affect the detection of various polyclonal antibodies circulating in the patient sera.
When the antigenic properties of several different preparations of
P. westermani were compared, serum and CSF samples from patients with active pulmonary or chronic cerebral paragonimiasis exhibited significantly high antibody levels to all antigenic preparations except for the metacercarial extract. Low levels of specific antibody against the metacercarial extract may be due to the rarity of antigenic materials as shown in
Fig. 1, in which specific antibodies reacted with only a few protein components (
Kong et al., 1997,
1998) even in the same protein concentrations. The crude extract of 4-week old juveniles showed the highest antibody levels in both active and chronic paragonimiasis by ELISA. However, it showed cross reactions with sera from other parasitic infections (
Kong et al., 1998) which suggested that this antigenic preparation was not applicable for the diagnosis in terms of specificity. Therefore, it seems to be reasonable that detection of 35 and 32 kDa either in 7-week worm or in adult extract would be a serological indicator for active paragonimiasis together with detection of rPw28CCP as shown in previous and present studies (
Kong et al., 1998;
Yun et al., 2000).
Monitoring the patients after specific treatment is important not only for the establishment of appropriate management of the patients but also for the proper evaluation of drug efficacy and for the detection of possible hidden lesions, especially in tissue invading helminth infections. However, there are several reports in which different outcomes have been described after treatment as far as the results of serological follow-up are concerned. In fascioliasis, for example, specific antibody levels in the patient sera returned mostly to their negative ranges between 6 and 12 months after successful treatment, whereas treatment-failure cases exhibited persistent antibody reactions (
Apt et al., 1995). Serum levels of the specific IgG and IgM antibodies against both adult and egg extracts were reported to persist during the follow-up period of two years in
Schistosoma mansoni infection (
Rabello et al., 1997) whereas others described that the antibody responses against egg protein turned to negative range within the follow-up of one year (
Noya et al., 1995). These discrepant findings may be lie on the fact that the observation period is not long enough to allow full serological profiles, especially those involving post-treatment. In addition, pathologic conditions associated with egg deposition in the tissues may give rise to these complex results in both schistosomiasis and fascioliasis.
In the present study of serological follow-up after treatment, one group of patients showed the patterns of declining antibody levels as previously described (
Cho et al., 1989). Their antibody levels declined in relatively short period. In these cases, antibodies recognizing the 35 and 32 kDa in adult and in 7-week worms and the 27 kDa of egg protein tended to fall below the positive criterion shortly after the treatment as analyzed by both immunoblot and immunoprecipitation. Reaction to the rPw28CCP also disappeared rapidly. However, in another group of paragonimiasis patients, the antibody levels persisted throughout the prolonged period. In these cases, antibody levels reacting to the egg proteins were the highest without an exception. As shown in
Fig. 3B and 3D, sera from these patients showed that egg specific protein at 27 kDa persisted throughout the observation period whereas the antigenic proteins other than egg proteins such as 35 and 32 kDa disappeared earlier. The reaction against rPw28CCP also disappeared rapidly. Taken together, we concluded that the persisting high antibody levels after the specific treatment would be caused by continuous sensitization of primed B cells by egg proteins. Some paragonimiasis patients treated with praziquantel may retain egg granulomas which are not resolved by the treatment and may provoke the persistent antibody reactions.
The present results raise a question concerning the specificity of the antibody tests in the diagnosis of paragonimiasis. If we define paragonimiasis as the patients infected with living worms only, the serum levels of persisting antibodies after specific treatment reflect lowered specificity, because egg granuloma in human paragonimiasis are subjects of surgical removal but not of chemotherapy. In this clinical settingof egg granulonma, we would prefer negative results in the antibody tests in view of chemotherapy. On the other hand, if we define specificity of the antibody test as differentiating paragonimiasis from other causes of lung or brain diseases including cancer, tuberculosis, fungus ball and bacterial infections, the high antibody levels in patients with ectopic egg granuloma may also be informative. For example, in chronic cerebral paragonimiasis, high levels of specific antibodies are helpful in differentiating brain lesions from other causes of space occupying lesions. In this respect, egg antigen is a unique component in the diagnosis of a wide clinical spectrum of human paragoni-miasis.
In conclusion, persisting antibody reaction in paragonimiasis after successful treatment is caused by continuous antigenic challenge from egg granuloma. It also suggests that differential antibody tests simultaneously employing several antigenic preparations such as eggs, metacercariae, adults and rPw28CCP in paragonimiasis can differentiate active cases from chronic or other causes of infectious lung granuloma.
Notes
-
This work was supported by a research grant for basic medical sciences, Ministry of Education (1996-1998), Republic of Korea.
References
- 1. Apt W, Aguilera X, Vega F, et al. Treatment of human chronic fascioliasis with triclabendazole: drug efficacy and serologic response. Am J Trop Med Hyg 1995;52:532-535.
- 2. Blair D, Xu ZB, Agatsuma T. Paragonimiaisis and the genus Paragonimus. Adv Parasitol 1999;42:113-222.
- 3. Cha SH, Chang KH, Cho SY, et al. Cerebral paragonimiasis in early active stage: CT and MR features. Am J Roentgenol 1994;162:141-145.
- 4. Cho SY, Hong ST, Rho YH, Choi SY, Han YC. Application of Micro-ELISA in serodiagnosis of human paragonimiasis. Korean J Parasitol 1981;19:151-156.
- 5. Cho SY, Kim SI, Kang SY, et al. Antibody changes in paragonimiasis patient after praziquantel treatment as observed by ELISA and immunoblot. Korean J Parasitol 1989;27:15-21.
- 6. Cho SY, Kong Y, Kang SY. Epidemiololgy of paragomimiasis in Korea. Southeast Asian J Trop Med Public Health 1997;28(suppl.):32-36.
- 7. Chung YB, Kong Y, Joo IJ, Cho SY, Kang SY. Excystment of Paragonimus westermani metacercariae by endogenous cysteine protease. J Parasitol 1995;81:137-142.
- 8. Chung YB, Kong Y, Yang HJ, Kang SY, Cho SY. Cysteine protease activities during maturation stages of Paragonimus westermani. J Parasitol 1997;83:902-907.
- 9. Garraud O, Perler FB, Bradley JE, Nutman TB. Induction of parasite antigen-specific antibody responses in unsensitized human B cells is dependent on the presence of cytokines after T cell priming. J Immunol 1997;159:4793-4798.
- 10. Ikeda T. Antibody responses to fluke cysteine proteinases in Paragonimus- and Fasciola-infected rats. J Helminthol 1998a;72:187-191.
- 11. Ikeda T. Cystatin capture enzyme-linked immunosorbent assay for immunodiagnosis of human paragonimiasis and fascioliasis. Am J Trop Med Hyg 1998b;59:286-290.
- 12. Ikeda T, Oikawa Y, Nishiymaya T. Enzyme-linked immunosorbent assay using cysteine proteinase antigens for immunodiagnosis of human paragonimiasis. Am J Trop Med Hyg 1996;55:435-437.
- 13. Im JG, Kong Y, Shin YM, et al. Pulmonary paragonimiasis: Clinical and experimental studies. RadioGraphics 1993;13:575-586.
- 14. Im JG, Whang HY, Kim WS, Han MC, Shin YM, Cho SY. Pleuropulmonary paragonimiasis: Radiologic findings in 71 patients. Am J Roentgenol 1992;159:39-43.
- 15. Kang SY, Cho MS, Chung YB, Kong Y, Cho SY. A cysteine protease of Paragonimus westermani eggs. Korean J Parasitol 1995;33:323-330.
- 16. Kang SY, Kong Y, Cho SY. Component proteins in crude extract of adult Paragonimus westermani purified by immunoaffinity chromatography using monoclonal antibodies. Korean J Parasitol 1991;29:363-369.
- 17. Knobloch J, Paz G, Feldmeier H, Wegner D, Voelker J. Serum antibody levels in human paragonimiasis before and after therapy with praziquantel. Trans Roy Soc Trop Med Hyg 1984;78:835-836.
- 18. Kong Y, Chung JY, Yun DH, et al. Variation of antigenic proteins of eggs and developmental stages of Paragonimus westermani. Korean J Parasitol 1997;35:197-202.
- 19. Kong Y, Ito A, Yang HJ, et al. Immunoglobulin G (IgG) subclass and IgE responses in human paragonimiases caused by three different species. Clin Diagn Lab Immunol 1998;5:474-478.
- 20. Maleewong W, Intapan PM, Priammuenwai M, et al. Monoclonal antibodies to Paragonimus heterotremus and their potential for diagnosis of paragonimiasis. Am J Trop Med Hyg 1997;56:413-417.
- 21. Noya O, Losada S, Alarcon de Noya B, et al. Effect of chemotherapy on immune response to egg antigens of Schistosoma mansoni in chronically infected children from areas of low transmission. Parasite Immunol 1995;17:111-117.
- 22. Nomura M, Nitta H, Nakada M, Yamashima T, Yamashita J. MRI findings of cerebral paragonimiasis in chronic stage. Clin Radiol 1999;54:622-624.
- 23. Rabello AL, Garcia MM, Pinto da Silva RA, Rocha RS, Katz N. Humoral immune responses in patients with acute Schistosoma mansoni infection who were followed up for two years after treatment. Clin Infect Dis 1997;24:304-308.
- 24. Slemenda SB, Maddison SE, Jung EC, Moore DD. Diagnosis of paragonimiasis by immunoblot. Am J Trop Med Hyg 1989;39:469-471.
- 25. Yang HJ, Chung JY, Yun DH, et al. Immunoblot analysis of a 10 kDa antigen in cyst fluid of Taenia solium metacestodes. Parasite Immunol 1998;20:483-488.
- 26. Yokogawa M. Paragonimus and paragonimiasis. Adv Parasitol 1965;3:99-158.
- 27. Yun DH, Chung JY, Chung YB, et al. Structural and immunological characteristics of a 28 kDa cruzipain-like cysteine protease of Paragonimus westermani expressed in the definitive host stage. Clin Diagn Lab Immunol 2000;7:932-939.
- 28. World Health Organization. Control of foodborne trematode infections: report of a WHO study group. WHO Tech Rep Ser 1995;849:1-157.
Fig. 1Immunoblot findings of crude extracts of the metacercariae, 4- and 7-week old juvenile worms, 16-week old adult worms and egg with mAb (A) or rabbit antiserum immunized with adult Paragonimus extract (B). After transfer-blot of the proteins separated by 7.5-15% SDS-PAGE, culture supernatant (undiluted) or rabbit antiserum (1:5,000 dilution) was probed overnight. The mAb exhibits positive reactions only with the adult and egg extracts (lanes adult and egg) while multiple immunoreactive bands are shown in panel B which was probed with the rabbit anti-serum. mc*, metacercariae; Mr, molecular mass in kDa.

Fig. 2A declining pattern of specific antibody levels in follow-up tests against each differently prepared antigens of Paragonimus westermani assessed by ELISA, immunoblot and immunoprecipitation. (A) Typical changes of antibody levels after praziquantel treatment analyzed by ELISA. The patient sera, collected serially after chemotherapy on 2, 8 and 28 months, exhibit the declining pattern regardless of antigenic preparations. Abs. against extracts from metacercariae (○-○), 4-week (□-□), 7-week (▵-▵), adult (★-★) and eggs (▾-▾). (B) Each serum secured from the time of diagnosis, 8 and 28 months after treatment (lanes a-c) is reacted with antigens of metacercaria (panel mc), 4-week (panel 4), 7-week (panel 7) and adult worms (panel 16), eggs (panel egg) and the recombinant protein of 28-kDa cruzipain like cysteine protease (panel rec), respectively. Strong reactions at 35 and 32 kDa in 7-week and adult extracts at the time of diagnosis disappeared rapidly with the time lapse. Weak reaction against 27 kDa of egg protein is observed. The recombinant protein also showed positive reaction at the time of the diagnosis and shows no reactions in the follow-up sera. The sera used in this experiment is the same as in A. Mr, molecular mass in kDa. (C) Another case whose serum antibody titers shows similar declining pattern as in A. Markings are the same as in A. (D) The follow-up sera in C is immunoprecipitated with adult extract and probed with mAb specific to egg protein. Specific antibody reactions at 27 kDa in egg protein (▸) become fainter and disappeared 8 month after treatment.

Fig. 3Another changing pattern of serum levels of anti-
Paragonimus westermani after praziquantel treatment. The specific antibody levels in the patient sera persisted.
(A) The patient sera, collected serially 3, 6, 11, 15 and 23 months after treatment show high antibody levels against the egg and adult extracts while those against other antigenic preparations decreased more promptly. Markings are same as in
Fig. 2.
(B) The follow-up sera of A reacted with different antigens by immunoblot. Each serum collected at the time of diagnosis, 6, 15 and 23 months post-treatment (lanes
a-
d) is reacted with different crude extracts as shown in
Fig. 2B. Strong reactions at 35 and 32 kDa in 7-week and adult extracts disappeared rapidly within 15 months follow-up; reactions against egg protein, at 27 kDa protein sustained throughout the follow-up period of 23 months. The reaction against recombinant protein also disappear quickly.
(C) Sera from another patient collected serially 2, 10, 15, 22, 27 and 34 months after successful treatment show high antibody reactions against the egg extract. Markings are the same as described in
Fig. 2. M
r, molecular mass in kDa.
(D) Immunoprecipitation analysis of the follow-up sera of a case presented in Fig. 3C. Reactions at 94 and 46 kDa faded out within 10 months after treatment (open arrowheads) while the reaction at 27 kDa persisted throughout the observation period of 34 months (closed arrowhead).

Table 1.Antibody levels by ELISA in sera/CSFs of active and chronic paragonimiasis patients to respective antigenic preparation
Table 1.
|
Antigens |
Absorbance (mean±SD)
|
|
Active paragonimiasis
|
Chronic paragonimiasis
|
|
Serum (n=16) |
Serum (n=22) |
CSFs (n=13) |
|
Metacercaria |
0.34±0.17 |
0.38±0.31 |
0.16±0.07 |
|
4-week old juvenile |
0.88±0.20 |
0.69±0.37 |
0.47±0.27 |
|
7-week old immature |
0.79±0.20 |
0.60±0.30 |
0.39±0.20 |
|
16-week old adult |
0.80±0.23 |
0.61±0.35 |
0.45±0.19 |
|
egg |
0.84±0.32 |
0.43±0.29 |
0.45±0.32 |