Abstract
Diphyllobothrium latum and Diphyllobothrium nihonkaiense are the 2 reported main causes of human diphyllobothriasis in the Republic of Korea. However, the differentiation of these 2 species based on morphologic features alone is difficult. The authors used nucleotide sequencing of the mitochondrial cytochrome c oxidase subunit 1 (cox1) gene to diagnose Diphyllobothrium spp. Two patients visited the emergency room at Kyungpook National University Hospital on 3 April and 12 April 2013, respectively, with fragments of parasites found while defecating. The parasites were identified as Diphyllobothrium spp. based on morphologic characteristics, and subsequent cox1 gene sequencing showed 99.9% similarity (1,478/1,480 bp) with D. nihonkaiense. Our findings support the hypothesis that D. nihonkaiense is a dominant species in Korea.
-
Key words: Diphyllobothrium nihonkaiense, human case, praziquantel, cox1 gene
INTRODUCTION
Diphyllobothriasis is caused by the adult tapeworm (
Diphyllobothrium spp.), and is contracted by consuming raw or undercooked fish in Northwest Europe and East Asia. Many patients with diphyllobothriasis are asymptomatic, while other patients develop gastro-intestinal discomfort, diarrhea, or abdominal pain [
1]. Of the 18
Diphyllobothrium spp. known to cause human diphyllobothriasis,
Diphyllobothrium latum and
Diphyllobothrium nihonkaiense are usually reported in Republic of Korea [
2,
3]. These 2
Diphyllobothrium species have similar morphologic features, and thus, clinicians are not able to differentiate the 2 parasites by naked eyes. As an alternative, genetic studies can be used to identify the species [
4,
5]. Recently, several nucleotide sequencing analysis studies revealed that previous identifications of
D. latum based on morphologic characteristics were incorrect and that the parasite was in fact
D. nihonkaiense [
5,
6,
7]. Although the chemotherapy required for both species is identical, the correct identification of
Diphyllobothrium species is required from the perspectives of epidemiology and public health promotion, particularly by providing accurate information on intermediate hosts [
8]. In this regard, we used nucleotide sequencing analysis of the mitochondrial cytochrome
c oxidase subunit I (
cox1) gene to identify
Diphyllobothrium species in 2 Korean male patients.
CASE RECORD
On April 3rd and 12th of 2013, 2 patients visited Kyungpook National University Hospital with a whitish yellow tapeworm segment. The patient that visited on April 3 was a healthy 50-year-old man (patient A), who visited the emergency room after finding a part of a tapeworm while defecating. He had not experienced any abdominal discomfort or pain, and had not visited any foreign country recently, and reported infrequent consumption of raw salmon and trout in side dishes. His blood test results were normal. The other patient was a healthy 20-year-old man (patient B). He also felt a foreign body while defecating, and visited the same emergency room. He stated that he seldom consumed raw fish. No abnormal clinical signs, such as, abdominal discomfort or pain, were observed, and he has taken a vermicide during September 2012. His blood test results were also normal.
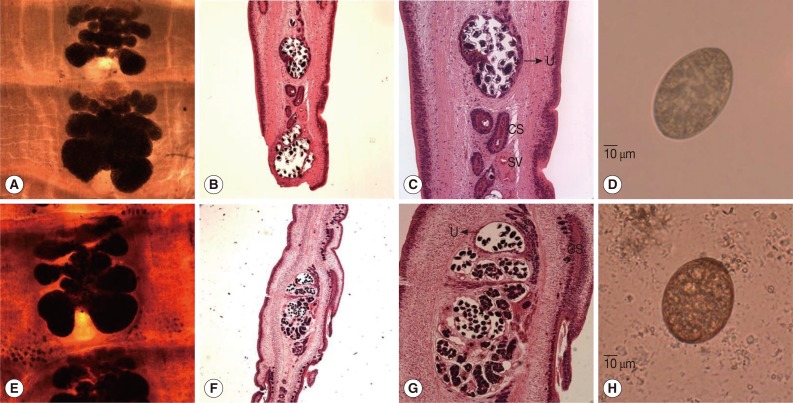
The specimens obtained from the 2 patients were forwarded to our laboratory with a request to identify the parasite species concerned. The parasite samples from the 2 patients consisted of a number of proglottids; gravid proglottids showed a rosette-shaped uterus as shown in
Fig. 1A and E. Testes were follicular and ovaries were kidney-shaped in the posterior region. In longitudinal sections of gravid proglottids, uterine and genital pores were separated at midline, and the uterus was filled with eggs (
B, C, F, and G in Fig. 1; H&E stain), which were oval and enclosed by a shell (
D and H in Fig. 1). Based on these morphologic characteristics, both parasites were identified as
Diphyllobothrium spp.
For genetic analysis, total genomic DNA was extracted from each specimen, and oligonucleotide primers were designed to amplify the 1,480-bp product of the mitochondrial cytochrome c oxidase subunit 1 (cox1) gene (forward primer, 5'-ATGACTAATCTTAAAGTTTT-3'; reverse primer, 5'-TAAAGCCAACATACTATAATC-3'). PCR was performed as follows; initial denaturation at 96℃ for 5 min, followed by 30 amplification cycles (96℃ for 1 min, 58℃ for 1 min, and 72℃ for 1 min), and a final extension step at 72℃ for 10 min. The purified PCR-amplified fragments obtained were cloned into pGEM®-T Easy Vector (Promega, Madison, Wisconsin, USA), and the inserted plasmids (cox1 gene-pGEM®-T Easy Vector) were sequenced using a Big-Dye Terminator sequencing kit (Applied Biosystems, Foster City, California, USA) on an automated DNA sequencer (Applied Biosystems). Sequences were assembled and aligned using CLC Sequence Viewer 6.8.1. (CLC Bio, Aarhus, Denmark), and similarities in GenBank were examined using BLASTN. The cox1 sequences (1,480 bp) of the 2 parasites showed similarities of 99.9% (1,478/1,480 bp), 95.1% (1,407/1,480 bp), 93.1% (1,378/1,480 bp), and 92.6% (1,371/1,480 bp) with D. nihonkaiense (GenBank no. AB781787), D. ursi (GenBank no. AB605762), D. dendriticum (GenBank no. AM412738), and D. latum (GenBank no. AM712906), respectively.
The patients were treated with a single oral dose of praziquantel (15 mg/kg) and no evidence of recurrence has been noticed.
DISCUSSION
D. nihonkaiense was first identified by Yamane et al. [
9] in 1986 by comparing its morphologic characteristics, such as egg size, the pit shape of eggshells, and the angle between the long axis of the cirrus sac and seminal vesicle, with those of other
Diphyllobothrium species [
9].
D. nihonkaiense and
D. latum had distinct morphologic characteristics. Specifically, the average size of
D. nihonkaiense eggs was 55.5 (±1.0) × 40.5 (±1.5) µm, which is smaller than the average size of
D. latum eggs (greater than 60.0 × 45.0 µm). The eggshells of
D. nihonkaiense had shallower pits distributed on the smooth surface than the eggshells of
D. latum did. The cirrus sac is situated obliquely in
D. nihonkaiense, and the angle between the long axis of the cirrus sac and seminal vesicle was sharper in
D. nihonkaiense than in
D. latum. In addition to these distinct morphologies,
D. nihonkaiense and
D. latum have different intermediate host specificities. The intermediate hosts of
D. nihonkaiense are salmon, such as cherry, pink, chum, and sockeye salmon (
Onchorhychus masou,
O. gorbuscha,
O. keta, and
O. nerka, respectively), which are found in the northern Pacific, whereas the intermediate hosts of
D. latum are perch, pike, burbot, and walleye [
8]. Based on his history, patient A likely contracted parasites from these fish. On the other hand, patient B had no history of raw salmon consumption, and similarly, several other cases of
D. nihonkaiense infection not associated with salmon consumption have been reported [
1]. Accordingly, accumulating evidence indicates
D. nihonkaiense has another intermediate host and that further research on the intermediate hosts of
D. nihonkaiense in Korea is needed.
Previously, most cases of diphyllobothriasis in Korea were thought to be caused by
D. latum. However, this paradigm shifted when Eom et al. [
10] reported occurrence of
D. nihonkaiense in Korea (35th annual meeting of The Korean Society for Parasitology, Kwangju, ROK). Owing to the publication of the complete mitochondrial genome of
D. nihonkaiense [
10],
D. nihonkaiense can now be accurately identified by DNA sequencing of the mitochondrial
cox1 gene. In 2009, Jeon et al. [
5] reported that 62 cases diagnosed as
D. latum infection were actually caused by
D. nihonkaiense, which were identified by genetic analysis. Moreover, another case of diphyllobothriasis was recently confirmed as
D. nihonkaiense infection by
cox1 gene analysis [
7]. These findings suggest that
D. nihonkaiense has been misdiagnosed as
D. latum and that
D. latum might not exist in Korea. The present study, in which 2 Korean male patients were diagnosed as
D. nihonkaiense by
cox1 gene sequencing, supports this suggestion. From the clinical point of view, this specific diagnosis might seem academic because diphyllobothriasis can be easily cured with praziquantel. Nevertheless, specific diagnosis at the species level is important in the contexts of epidemiology and public health, and would provide further insight into distribution of different taxa and improve our understanding of imported cases and outbreaks of diphyllobothriasis.
Notes
-
We have no conflict of interest related to this study.
References
- 1. Kamiya M, Ooi HK. Current status of food-borne parasitic zoonoses in Japan. Southeast Asian J Trop Med Public Health 1991;22:48-53.
- 2. Lee EB, Song JH, Park NS, Kang BK, Lee HS, Han YJ, Kim HJ, Shin EH, Chai JY. A case of Diphyllobothrium latum infection with a brief review of diphyllobothriasis in the Republic of Korea. Korean J Parasitol 2007;45:219-223.
- 3. Yamane Y, Shiwaku K. Diphyllobothrium nihonkaiense and other marine-origin cestodes. Prog Med Parasitol Jpn 2003;8:245-254.
- 4. Wicht B, de Marval F, Peduzzi R. Diphyllobothrium nihonkaiense (Yamane et al., 1986) in Switzerland: first molecular evidence and case reports. Parasitol Int 2007;56:195-199.
- 5. Jeon HK, Kim KH, Huh S, Chai JY, Min DY, Rim HJ, Eom KS. Morphologic and genetic identification of Diphyllobothrium nihonkaiense in Korea. Korean J Parasitol 2009;47:369-375.
- 6. Choi HJ, Lee J, Yang HJ. Four human cases of Diphyllobothrium latum infection. Korean J Parasitol 2012;50:143-146.
- 7. Park SH, Eom KS, Park MS, Kwon OK, Kim HS, Yoon JH. A case of Diphyllobothrium nihonkaiense infection as confirmed by mitochondrial COX1 gene sequence analysis. Korean J Parasitol 2013;51:471-473.
- 8. Scholz T, Garcia HH, Kuchta R, Wicht B. Update on the human broad tapeworm (genus Diphyllobothrium), including clinical relevance. Clin Microbiol Rev 2009;22:146-160.
- 9. Yamane Y, Kamo H, Bylund G, Wikgren BJP. Diphyllobothrium nihonkaiense sp. nov. (Cestoda: Diphyllobothriidae)-revised identification of Japanese broad tapeworm. Shimane J Med Sci 1986;10:29-48.
- 10. Kim KH, Jeon HK, Kang S, Sultana T, Kim GJ, Eom KS, Park JK. Characterization of the complete mitochondrial genome of Diphyllobothrium nihonkaiense (Diphyllobothriidae: Cestoda), and development of molecular markers for differentiating fish tapeworms. Mol Cells 2007;23:379-390.
Fig. 1Gravid proglottids and eggs of Diphyllobothrium nihonkaiense from the 2 patients. (A, E) Whole-mounted specimens of a proglottid showing the uterus and cirrus sac (×40 magnification). (B, C, F, G) Longitudinal sections of a mature proglottid showing the uterus (U), cirrus sac (CS), and seminal vesicles (SV) (H&E stain; B and F,×40; C and G,×100). (D, H) Eggs discharged from the gravid proglottid of D. nihonkaiense (×400). (A-D, D. nihonkaiense from patient A; E-H, D. nihonkaiense from patient B).