Abstract
To evaluate the usefulness of the Korean Isolate-1 (KI-1) antigen for serodiagnosis of toxoplasmosis, antigen profiles of KI-1 tachyzoites were analyzed in comparison with RH tachyzoites by SDS-PAGE and immunoblotting. ELISA was performed on latex agglutination (LA)-positive and negative serum samples using KI-1 and RH antigens. Immunoblotting of the KI-1 antigen showed multiple antigen bands with molecular sizes of 22-105 kDa. Among them, 1 and 6 common bands were noted against a KI-1-infected and a RH-infected human serum, respectively, which represented differences in antigenic profiles between KI-1 and RH tachyzoites. However, all 9 LA-positive human sera were found positive by ELISA, and all 12 LA-negative sera were negative by ELISA; the correlation between the ELISA titers and LA titers was high (r = 0.749). Our results suggest that tachyzoites of KI-1 may be useful for serodiagnosis of human toxoplasmosis.
-
Key words: Toxoplasma gondii, Toxoplasmosis, Korean isolate-1 (KI-1), RH strain, Serodiagnosis, Antigen
Toxoplasma gondii is an intracellular protozoan that can infect various species of birds and mammals, including humans [
1,
2]. This protozoan is prevalent in 20-80% of human population in different territories of the world [
3,
4]. Moreover, in a cross-sectional study in the U.S. (1988-1994), about a quarter of adolescents and adults were found to be infected with
T. gondii [
5], and the prevalence was not changed in a succeeding survey (1999-2000) [
6]. In the Republic of Korea, the seroprevalence was reported to be lower than that of the Europe and New World countries, i.e., around 2-7% among the general population [
7], and 7.7% among children less than 10 years of age [
8].
A new geographical isolate of the
T. gondii virulent group, named Korean Isolate-1 (KI-1), was recently isolated from the blood of an ocular patient in the Republic of Korea [
9], and has been successfully maintained in the laboratory. Its morphology, virulence in mice, and cell culture characteristics were similar to those of the well-known virulent RH strain, which originated from a child who suffered from encephalitis [
10]. In genotype analyses, KI-1 belonged to the virulent clonal lineage, i.e., genotype I, in which RH is included [
11,
12]. However, immunological properties, including antigenic profiles and feasibility as a diagnostic antigen, have never been documented.
For serodiagnosis of human toxoplasmosis, the latex agglutination (LA) test using the RH antigen (Toxotest-MT kit, Eiken, Tokyo, Japan) has been commonly used [
8,
13,
14]. It is suggested that KI-1 may be immunologically similar to RH, and could be used as an alternative of the RH antigen. However, for proper assessment of its efficiency and feasibility, immunological characteristics, in particular the antigenic properties, of KI-1 should be analyzed. In the present study, we analyzed the antigenic profiles of KI-1 tachyzoites by SDS-PAGE and immunoblotting using human and rabbit polyclonal antibodies in comparison with RH tachyzoites, and evaluated the diagnostic potential of the KI-1 antigen by ELISA using LA-positive and LA-negative human serum samples.
Tachyzoites of
T. gondii KI-1 and RH were maintained in BALB / c mice by serial passages using the previously described procedure [
9,
10,
15]. Tachyzoites purified were disrupted by 5 cycles of freezing to - 70℃ and thawing at room temperature. The homogenate was centrifuged at 12,000 rpm at 4℃ for 30 min, and the supernatant was used as the lysate antigen. Protein components of KI-1 and RH tachyzoite lysates were separated by SDS-PAGE on 10% gels and transferred onto polyvinylidene fluoride nitrocellulose membranes (Millipore Corporation, Bedford, Massachusetts, USA) for Western blotting. The membranes were then incubated at room temperature for 1 hr with 1 of 3 polyclonal antibodies; a KI-1-infected human serum, an RH-infected human serum, and an RH-infected rabbit serum. The KI-1-infected human serum was the one obtained from the patient from whom the KI-1 was isolated [
9]. The RH-infected human serum was obtained from an author of this paper accidentally infected 6 month previously by a needle-stick during an experiment with RH tachyzoites. The RH-infected rabbit serum was obtained from a rabbit immunized with RH tachyzoites 3 month previously with boosting 2 times (provided by Zhejiang Academy of Medical Sciences, Zhejiang, China). Horseradish peroxidase (HRP)-conjugated Protein A and HRP-conjugated goat anti-human IgG (Cappel Co., St. Louis, Missouri, USA) were used as secondary antibodies. The immunoblots were developed with 4-chloro-1-naphthol containing 0.03% H
2O
2 in phosphate buffer (0.01 M, pH 7.4), and the reaction was stopped by washing in distilled water.
For the LA test and ELISA, a total of 21 human serum samples were used, which consisted of 9 serologically (LA) T. gondii-positive and 12 T. gondii-negative samples collected from people (10 men and 11 women; age 18-46 yr) residing in the Republic of Korea. Informed consent for use of serum samples for study purposes was obtained from each person. Sera of all subjects were stored at -70℃ until use.
The LA test was performed according to the instructions of the manufacturer (Toxotest-MT, Eiken Chemical Co., Tokyo, Japan), and were considered positive when agglutination was observed at serum dilutions of 1 : 32 or higher. ELISA was performed in polystyrene 96-well flat-bottomed microtiter plates (Corning Inc., Lowell, Massachusetts, USA), which were coated with 100 µl of carbonate-bicarbonate buffer (50 mM, pH 9.6) containing 1 µg of the KI-1 or RH antigen. The test sera were diluted to 1 : 100 and added to triplicate wells. Then, 100 µl of 1 : 1,000 diluted HRP-labeled sheep anti-human IgG (Biochemical Corp., Lakewood, New Jersey, USA) was added to each well, and the plates were incubated for 1 hr at 37℃. The colorimetric reaction was performed using a substrate solution consisting of tetramethylbenzidine and 0.1% H2O2, and was then stopped by the addition of 50 µl / well of 1 M H2SO4. The optical density (OD) was read at 450 nm using a microtiter plate reader (Molecular Device, Concord, Ontario, Canada). The cutoff value (in OD) was determined as the mean plus 3 SD of 12 Toxoplasma-negative (LA negative) human sera, i.e., 0.33 and 0.34 for KI-1 and RH antigens, respectively. All experiments were repeated at least 3 times, and one representative data point is presented.
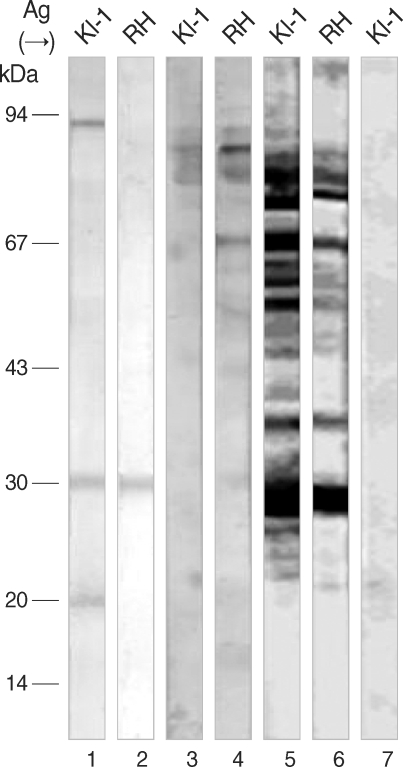
Immunoblots of
T. gondii KI-1 and RH tachyzoite lysates revealed multiple bands when reacted with KI-1- or RH-infected human or rabbit polyclonal antibodies, whereas lysates reacted with an uninfected rabbit serum revealed no bands (
Table 1). The KI-1-infected human serum detected 3 antigenic bands (91, 30, and 22 kDa) from the KI-1 antigen, and among them, 1 band (30 kDa) was in common with the RH (
Table 1;
Fig. 1). The RH-infected human serum detected 11 reaction bands (91, 85, 82, 80, 76, 67, 55, 48, 30, 23, and 19 kDa) from the RH antigen, and among them, 6 bands (91, 85, 82, 67, 30 and 23 kDa) were found in common with the KI-1 (
Table 1;
Fig. 1). By comparison, the RH-infected rabbit serum showed a total of 13 bands (105, 91, 85, 80, 67, 60, 55, 48, 45, 35, 30, 25, and 22 kDa) in response to the RH antigen, and among them, 12 were found in common with the KI-1 (
Table 1;
Fig. 1).
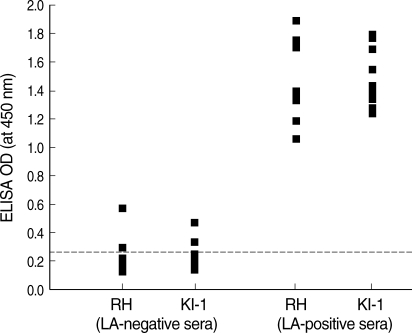
ELISA of 9 LA-positive and 12 LA-negative serum samples using the KI-1 or RH antigen revealed high correlations with the results of LA using the RH antigen (
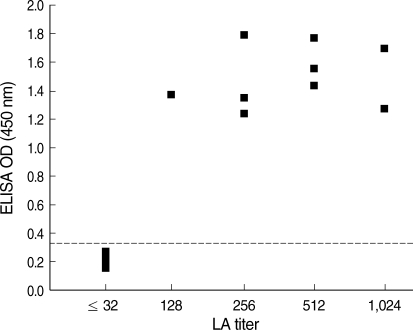
Fig. 2). All LA-negative sera (n = 12; titer ≤ 1 : 32) were found negative by ELISA (OD 0.14-0.28 against KI-1, and 0.13-0.29 against RH), and all LA-positive sera (n = 9; titer 1 : 128 to 1 : 1,024) were found positive by ELISA (OD 1.23-1.79 against KI-1, and 1.06-1.89 against RH). The distribution of LA titers to RH antigen and ELISA ODs to KI-1 antigen showed high correlations (correlation coefficient: 0.749) with each other (
Fig. 3).
To date, the diagnosis of toxoplasmosis has been undertaken using the RH antigen in most parts of the world, including the Republic of Korea. The diagnostic techniques included the indirect LA test, indirect fluorescent antibody test, ELISA, and immunoblotting [
7-
9,
13,
14]. However, it was unclear whether the RH antigen was sufficient to detect all the infections occurring in local areas despite the possible coexistence of different parasite strains. This steered for the investigation toward the establishment and maintenance of local strains or isolates as candidates of diagnostic antigens. KI-1 seemed to be a good candidate for this purpose, and it was needed to define the antigenic properties of KI-1 in comparison with RH using strain-specific human sera. Fortunately, we had both types of human sera, i.e., a KI-1-specific serum and an RH-specific serum. In addition, we had 9 LA-positive and 12 LA-negative sera from Korean people, and these could be used for a feasibility test by ELISA.
The results of the present study revealed that both KI-1 and RH antigens reacted against the KI-1-infected and the RH-infected human sera, as well as to the RH-infected rabbit serum, showing variable numbers of antigenic bands. However, the dominant antigens were located between 22-91 kDa. The most common antigen was 30 kDa, irrespective of KI-1 or RH, and human anti-KI-1 or rabbit anti-RH sera used, although, this 30 kDa band was faint when reacted against human anti-RH sera. The 30 kDa protein seems to be identical to the major known 30 kDa antigen of
T. gondii KI-1 and RH tachyzoites, SAG1 [
9,
16].
It is interesting to note that KI-1 tachyzoites revealed a higher number of antigenic proteins compared with RH tachyzoites when reacted with the RH-infected rabbit serum; 18 antigenic bands were detected from KI-1, whereas 13 bands were detected from RH. By comparison, the KI-1- and RH-infected human sera showed different immunoblot patterns. For example, the KI-1-infected human serum detected 91, 30, and 22 kDa antigens from KI-1 tachyzoites, but detected only 1 antigen, at 30 kDa, from RH tachyzoites. The 30 kDa antigen is a common antigen of different
T. gondii strains and isolates [
9]. The 91 kDa antigen was also detected from RH tachyzoites, when reacted with the RH-infected human serum. Therefore, only the remaining 22 kDa protein is considered to be an antigen that is unique to KI-1 tachyzoites. However, the usefulness of the 22 kDa antigen as a serodiagnostic antigen for KI-1 infection requires further verification. The reason for the higher numbers of reaction bands when reacted with the infected rabbit serum compared with the infected human sera may be due to different intensities of
T. gondii infection. The rabbit which was immunized with boosting 2 times may have undergone a severe infection, whereas the humans may have had less severe infections.
Conclusively, ELISA using the KI-1 antigen could clearly differentiate LA-positive and LA-negative human sera, demonstrating its high feasibility. Moreover, the ELISA ODs and LA titers were significantly correlated with each other. Therefore, KI-1 tachyzoites are suggested to be useful as a serodiagnostic antigen for human toxoplasmosis.
ACKNOWLEDGEMENTS
This work was supported by grant no. 02-05-013 from the Seoul National University Bundang Hospital (SNUBH) Research Fund (2005).
References
Fig. 1Immunoblots of T. gondii KI-1 (lanes 1, 3, 5 and 7) and RH (lanes 2, 4 and 6) tachyzoite lysates separated by 10% SDS-PAGE and labeled with KI-1- (lanes 1 and 2) or RH-infected human serum (lanes 3 and 4), RH-infected rabbit serum (lanes 5 and 6), or normal control rabbit serum (lane 7). Note antigenic profiles of KI-1 and RH. Molecular mass markers on left.

Fig. 2ELISA of human sera using 2 kinds of T. gondii antigens. All latex agglutination (LA) negative sera (n = 12) were negative by ELISA, and all LA positive sera (n = 9) were positive by ELISA. The cutoff values (0.34 for RH and 0.33 for KI-1 antigen) were set at the mean plus 3SD of negative sera, as shown by dotted lines. RH Ag; antigens prepared from RH tachyzoites, KI-1 Ag; antigens prepared from KI-1 tachyzoites.

Fig. 3The correlation between LA titer and ELISA OD for human sera using RH and KI-1 antigens, respectively (correlation coefficient: 0.749). All LA positive sera (n = 9), with titers higher than 1 : 64, were positive by ELISA, and all LA negative sera (n = 12) appeared negative by ELISA. The ELISA cutoff value (0.33) was set as the mean plus 3SD of negative sera, and shown by a dotted line.

Table 1.Number of immunoblot bands detected by antigen-specific sera against
Toxoplasma gondii KI-1
a and RH tachyzoites
Table 1.
|
Antigen |
No. of bands (No. of common bands) detected by
|
|
KI-1-infected human serumb
|
RH-infected human serumc
|
RH-infected rabbit serumd
|
|
KI-1 tachyzoites |
3 (1) |
7 (6) |
18 (12) |
|
RH tachyzoites |
1 (1) |
11 (6) |
13 (12) |