Abstract
We performed a molecular genetic study on the sequences of 18S ribosomal RNA (ITS1 region) gene in 4-day-old adult worms of Macroorchis spinulosus recovered in mice experimentally infected with metacercariae from crayfish in Jeollanam-do Province, Korea. The metacercariae were round, 180 μm in average diameter, encysted with 2 layers of thick walls, but the stylet on the oral sucker was not clearly seen. The adult flukes were oval shape, and 760-820 μm long and 320-450 μm wide, with anterolateral location of 2 large testes. The phylogenetic tree based on ITS1 sequences of 6 M. spinulosus samples showed their distinguished position from other trematode species in GenBank. The most closely resembled group was Paragonimus spp. which also take crayfish or crabs as the second intermediate host. The present study is the first molecular characterization of M. spinulosus and provided a basis for further phylogenetic studies to compare with other trematode fauna in Korea.
-
Key words: Macroorchis spinulosus, ITS1, sequencing, crayfish
Macroorchis spinulosus is an intestinal trematode of the family Nanophyetidae, which was named by its characteristically large testes. The genus
Macroorchis was erected by Ando [
1,
2] with
M. spinulosus in Japan, and 3 additional species have been reported in Japan [
3,
4]. The adults of
M. spinulosus were collected from the small intestine of mammals, infected naturally or experimentally, such as dogs, cats, mice, guinea pigs, rabbits, and albino and house rats [
1]. However, human infections have never been reported. Freshwater crabs,
Potamon dehaani and
Potamon obrusipes, and crayfish,
Cambaroides similis were known as intermediate hosts of
M. spinulosus [
1,
2]. Chai et al. [
2] redescribed
M. spinulosus in Korea based on adult worms because some morphological features of
M. spinulosus were different from the original descriptions by Ando [
1].
As other species,
Macroorchis himizu [
3],
Macroorchis chimarrogalus, and
Macroorchis elongatus [
4] have been described as new species in Japan according to recognition of one or more morphological differences, distinctive intermediate hosts, and geographical distribution. However, identification of trematodes with similar morphology is at times difficult and needs decision of a specialized person. In this case, molecular studies may be helpful for their identification as well as understanding the genetic relationships. However, no molecular information of
M. spinulosus has been available to date. Therefore, in this study, we analyzed genetic characteristics of
M. spinulosus on the basis of the sequences of ITS1 region in 18S ribosomal RNA.
A total of 10 crayfish,
Cambaroides similis, were caught in the streams in Isipgokri, Hwasun-eup, Hwasun-gun, Jeollanam-do Province, Korea from March to May 2015 (
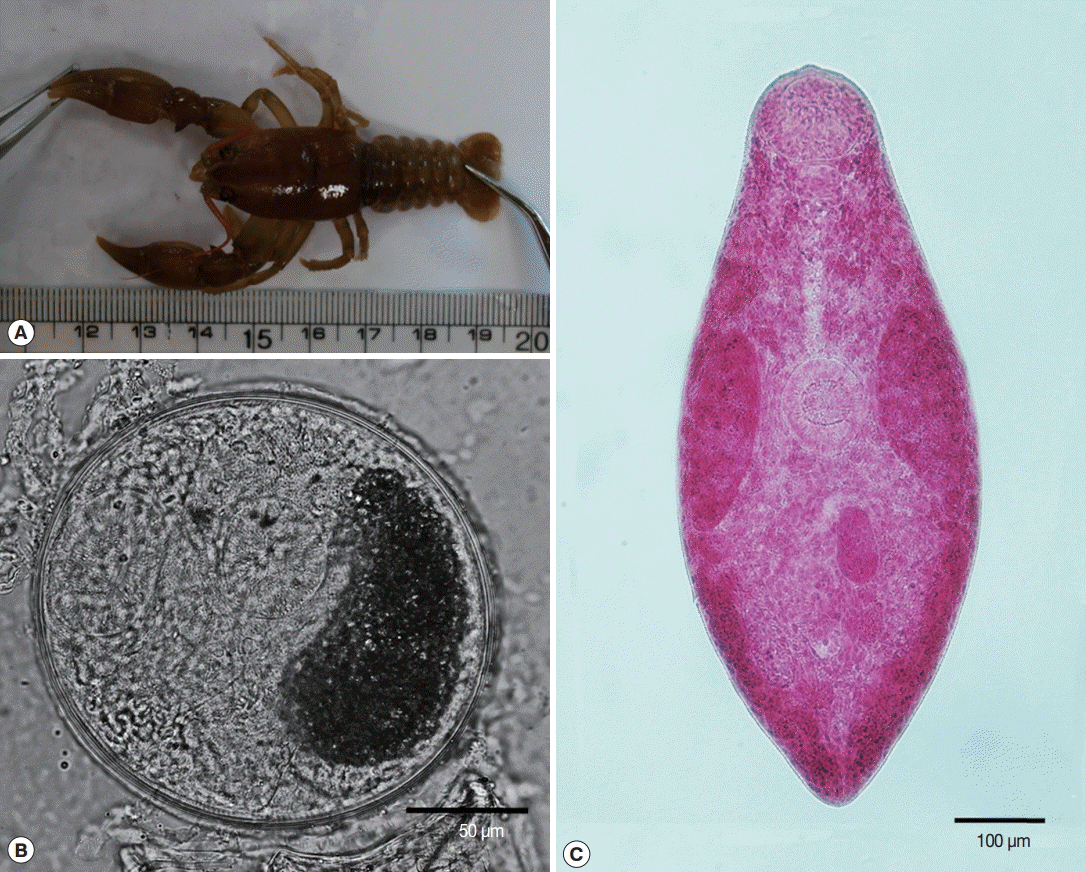
Fig. 1A). The crayfish were crushed with a mortar and a pestle, and washed with physiological saline.
M. spinulosus metacercariae were collected from the sediment under a dissecting microscope. Two female ICR mice at 6 weeks of age were orally fed each with 150 metacercariae. The mice were sacrificed at 4 days after the infection. The small intestine of mice was resected out and divided into the duodenum, jejunum, and ileum. Intestinal contents were washed in physiological saline, and
M. spinulosus flukes were recovered from the sediments under a dissecting microscope. The recovered metacercariae (excysted under cover slip pressure) and adult flukes were washed with cold physiological saline 3 times and fixed with 10% formalin, and stained with Semichon’s acetocarmine. Both fresh and stained specimens were observed under light microscopy.
In order to characterize the worms genetically, nucleotide sequences of 18S ribosomal RNA (ITS1 region) genes from adult worms were analyzed. The total genomic DNA (gDNA) was extracted by DNeasy Blood and Tissue Kit (Qiagen, Hilden, Germany) according to the manufacturer’s instructions. The DNA region comprising 18S rRNA, ITS1, and 5.8S rDNA was amplified by PCR using primers BD1 (forward: 5´-GTCGTAACAAGGTTTCCGTA-3´) [
3] and 5.8S_R1 (reverse: 5´-CGATGTTCAAAGCAGTATGC-3´) designed in this study. PCR was carried out in 25 μl of total volume, which contained 3 μl of DNA solution (20-40 ng), Smart 2× PCR premix Taq (Solgent, Daejeon, Korea), and 2 μl of each primer. The mixture was undergone initial denaturation at 94˚C for 3 min, followed by 45 cycles of denaturation at 94˚C for 40 sec, annealing at 55˚C for 45 sec, and extension at 72˚C for 1 min with a final extension at 72˚C for 5 min [
5]. The products were then separated by electrophoresis on 3% agarose gels containing 1 µg/ml ethidium bromide and visualized under ultraviolet light. Sequencing was carried out using a capillary automated ABI PRISM 3700 DNA sequencher (Macrogen Inc., Seoul, Korea). The GenBank Blast program was used for ITS sequences comparisons. After alignment of the sequences by ClustalW (Free Software Foundation, Boston, Massachusetts, USA), a phylogenetic tree was constructed using the neighbor-joining method.
A total of 355
M. spinulosus metacercariae were collected from 10 crayfishes. The metacercariae were round, 180 μm in average diameter, encysted with 2 layers of thick walls, but their stylet on the oral sucker was not distinctly seen (
Fig. 1B). The oral sucker was large. The acetabulum was smaller than the oral sucker and was located in mid-body. The excretory bladder was eccentric and contained numerous dark excretory granules. When the adult flukes were harvested from mice at 4 days post-infection, the overall recovery rate was about 30% (99/355). The adult flukes were oval in shape and 760-820 μm long and 320-450 μm wide, with anterolateral location of 2 large testes. The large testes occupied the mid-lateral regions of the body. Vitelline glands were follicular and distributed superficially from the posterior end of the body to the level of the pharynx. The uterus was convoluted in the portion of the body posterior to the ovary (
Fig. 1C).
Six randomly selected
M. spinulosus adult specimens were undergone sequencing of the ITS region. All of the samples revealed 82% identity with
Paragonimus ohirai (GenBank no. AF040930), 82% identity with
Paragonimus kellicotti (no. HQ900670.1), 81% identity with
Paragonimus westermani (no. AF040933, AF040932, AF040928), 79% identity with
Paragonimus macrorchis (no. AF040936), 76% identity with Metagonimus yokogawai (no. KJ631740.1), and 75% identity with
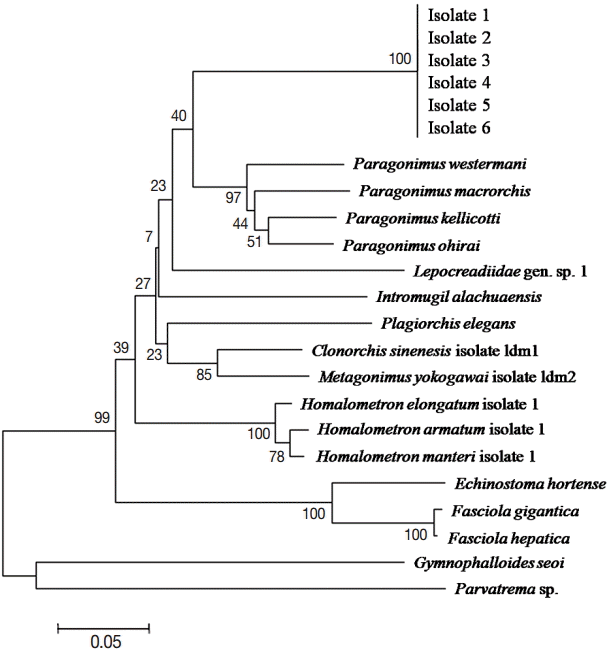
Clonorchis sinensis (no. KJ137226.1). The phylogenetic tree based on the ITS1 sequences showed a single clade of the 6
M. spinulosus samples analyzed in this study. However, they were distinguished from
Paragonimus species and other trematodes (
Fig. 2).
Although discrimination of different parasite species conventionally has been mainly based on morphological difference, limitation always exists due to subjective features according to the inspector. In addition to morphological characteristics, intermediate hosts, and regional distribution should also be considered in the proposal of the species. In Korea,
M. spinulosus was redescribed by Chai et al. [
2] as follows: oval body, ovary in variable position, median or shifted to the right or left side, apparently bilobed seminal vesicle, absence of seminal receptable, and vitellaria extending from the level of the pharynx, presence of a stylet on the oral sucker in about 1/4 of the specimens, nearly equal size of the acetabulum and oral sucker, ceca terminating near 1/5-1/6 body portion from the posterior extremity, genital pore just behind the ventral sucker, and more posterior location of the seminal vesicle.
Our specimens were consistent with these amended morphological descriptions. We also support that the crayfish is a second intermediate host for
M. spinulosus, caught from a stream in the southwestern area of Korea. Thus, in addition to Gangwon-do [
2] and Gyeongsangnam-do [
6] we suggest that Jeollanam-do Province should also be recognized as an ecological focus of
M. spinulosus in the Korean peninsula. Adult flukes of
M. spinulosus were collected from the small intestine of naturally or experimentally infected mammalian animals in Japan and Korea [
1,
2,
6-
8]. Therefore, humans may possibly be a natural definitive host which should be clarified in the future.
Interestingly, intermediate hosts of
M. spinulosus are different according to the region; in Japan, freshwater crabs,
P. dehaani and
P. obrusipes, were reported [
1]. However, in Korea, crayfish, C. similis, is known [
2]. Until now, only 4 English literature about
M. spinulosus are available [
2,
6-
8]. It is noted that this is the first molecular work which can be a cornerstone of phylogenic studies. Considering that there are some discrepancies in morphological features of
M. spinulosus between Japanese and Korean study [
1,
2], the use of molecular methods may be helpful for species confirmation and understanding their genetic relationships. Although there are 3 other species of
Macroorchis (
M. himizu,
M. chimarrogalus, and
M. elongatus) [
3,
4], we could not evaluate their genetic relationship with
M. spinulosus, because their molecular sequences have never been reported.
In the current study, a relatively close genetic relationship was found between M. spinulosus and Paragonimus spp. rather than the other trematodes. This finding is interesting and supported by that they share the same intermediate hosts, and also present morphological similarities in their eggs, cercariae, metacercariae, and adult worms, albeit their habitats were different (lung vs small intestine).
The present study is the first molecular characterization of M. spinulosus, allowing a reliable approach for genetic differentiation of Macroorchis spp. and providing basis for further studies on trematode fauna in Korea. We suggest that further studies using more polymorphic genetic markers such as mitochondrial DNA may be necessary for wider range of molecular analysis of Macroorchis and other related trematode species.
Notes
-
We have no conflict of interest related to this work.
Fig. 1.Morphological findings of Macroorchis spinulosus in this study. (A) Crayfish caught from Isipgokri, Hwasun-eup, Hwasun-gun, Jeollanam-do Province, Korea. (B) M. spinulosus metacercaria, which is round, 180 μm in average diameter, and encysted with 2 layers of thick walls. However, the stylet on the oral sucker is not distinct in this figure. (C) Adult worm of M. spinulosus showing its oval shape, 760-820 μm length and 320-450 μm width, and anterolateral location of 2 large testes (acetocarmine stain).

Fig. 2.Neighbor-joining tree showing the phylogenetic position of Macroorchis spinulosus based on ITS sequences of 6 adult specimens (isolates) analyzed in this study and its relationships with Paragonimus species and other trematodes available in GenBank.

References
- 1. Ando A. On a new trematode with the freshwater crab as its intermediate host. Tokyo Iji Shinshi 1918; no. 2081: 10-17 (in Japanese).
- 2. Chai JY, Sohn WM, Huh S, Choi MH, Lee SH. Redescription of Macroorchis spinulosus Ando, 1918 (Digenea: Nanophyetidae) encysted in the fresh water crayfish, Cambaroides similis. Korean J Parasitol 1996;34:1-6.
- 3. Machida M, Uchida A. Some helminth parasites of the Japanese shrew mole from the Izu Peninsula. Memo Nat Sci Museum 1982;15:149-154.
- 4. Saito Y, Watanabe T, Yamashita T. Two new intestinal trematodes, Macroorchis chimarrogalus n. sp. and Macroorchis elongatus n. sp. from Japanese water shrew and others, with a description of the metacercaria of Macroorchis chimarrogalus n. sp. (Trematoda: Nanophyetidae). Acta Med Biol 1982;30:47-56.
- 5. Luton K, Walker D, Blair D. Comparison of ribosomal internal transcribed spacers from two congeneric species of flukes (Platyhelminths: Trematoda: Digenea). Mol Biochem Parasitol 1992;56:323-328.
- 6. Woo HC, Chai JY, Hong SJ. Recovery, growth and development of Macroorchis spinulosus in albino rats. Korean J Parasitol 2003;41:27-33.
- 7. Sato H, Suzuki K. Gastrointestinal helminths of feral raccoons (Procyon lotor) in Wakayama Prefecture, Japan. J Vet Med Sci 2006;68:311-318.
- 8. Hong SJ, Woo HC, Kwon OS. Developmental surface ultrastructure of Macroorchis spinulosus in albino rats. Korean J Parasitol 2004;42:151-157.